
Lithoselatium tantichodoki, Promdam, Rueangrit & Ng, Peter K. L., 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.191392 |
|
DOI |
https://doi.org/10.5281/zenodo.6214787 |
|
persistent identifier |
https://treatment.plazi.org/id/D07F407E-B42F-FF84-ADE6-D6800B80066E |
|
treatment provided by |
Plazi |
|
scientific name |
Lithoselatium tantichodoki |
| status |
sp. nov. |
Lithoselatium tantichodoki View in CoL new species
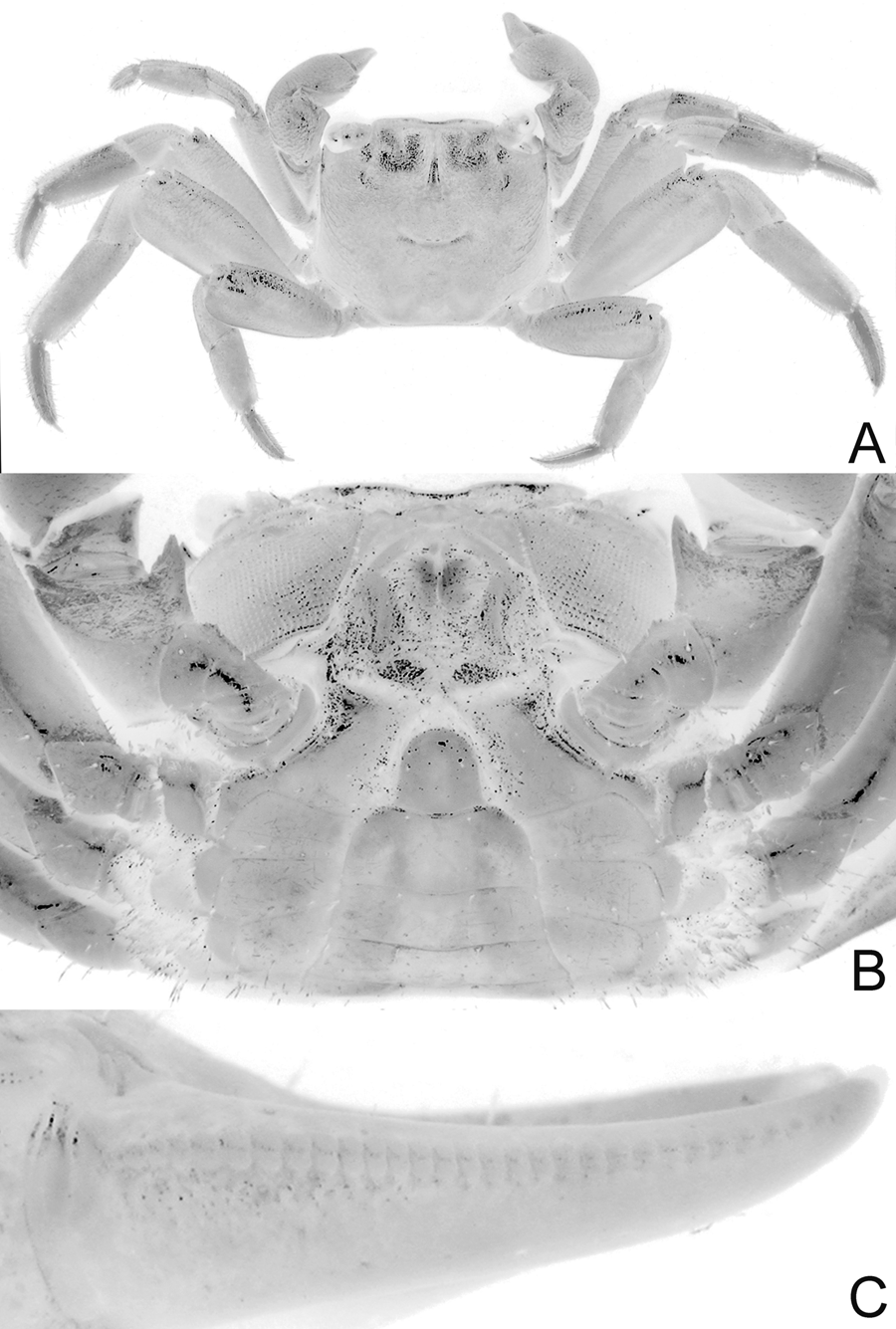
( Figs. 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 , 9A, C View FIGURE 9. A, C )
Material examined. Holotype: male (25.0 x 22.9 mm) ( ZRC 2009.1140, ex PMBC 25378), Ko Wang Nok, south of Ko Samui, Ko Samui District, Surat Thani Province, southern Thailand, Gulf of Thailand (9°18'45"N, 99°54'42"E), coll. R. Promdan, 27 January 2007. Paratypes: 1 ovigerous female (18.8 x 16.6 mm) ( PMBC 25380), Ko Tan, south of Ko Samui, Ko Samui District, Surat Thani Province, southern Thailand, Gulf of Thailand (9°23'33"N, 99°57'06"E), coll. R. Promdan, 1 April 2007; 1 juvenile female (12.6 x 11.0 mm) ( PMBC 25379), Ko Tan, south of Ko Samui, Ko Samui District, Surat Thani Province, southern Thailand, Gulf of Thailand (9°23'33"N, 99°57'06"E), coll. R. Promdan, 17 February 2007.
Comparative material. Lithoselatium pulchrum Schubart, Liu & Ng, 2009 : holotype male (23.5 x 20.8 mm) ( ZRC 2002.0152), Yakoulu, Manchou, Pingtung County, Taiwan, rocky shore with tide pools, coll. H.-C. Liu, 27 August 1999; paratypes: 1 female (23.2 x 20.1 mm with eggs) ( ZRC 2002.0152), same data as holotype; 1 female (18.1 x 15.7 mm), Chuanfanshih, Hengchun, Pingtung County, Taiwan ( ZRC 1999.0555), coll. P. K. L. Ng et al., 31 May 1997; 1 male (18.1 x 15.1 mm) ( ZRC), Chuanfanshih, Hengchun, Pingtung County, Taiwan, coll. P.-H. Ho, 2004. Lithoselatium kusu Schubart, Liu & Ng, 2009 : holotype male (16.0 x 14.35 mm) ( ZRC 2002.0149), Kusu Island, Singapore, between loose rocks, coll. C. D. Schubart, 26 December 1999; paratypes: 1 male (16.8 x 14.95 mm), 2 juvenile females (larger 14.15 x 12.4 mm) ( ZRC 2002.0150), same collection data as holotype; 1 juvenile female ( ZRC 2009.0568), Sentosa Island, Singapore, rock jetty, coll. C. D. Schubart, 15 August 1999; 7 males (largest 29.0 x 25.8 mm), 9 females (largest 29.1 x 26.0 mm) ( ZRC 2009.0569), Raffles Lighthouse, Singapore, coll. H. H. Tan et al., 30 January 2002; 1 female (21.2 x 19.0 mm) ( ZRC 2009.0570), Raffles Lighthouse, Singapore, coll. P. K. L. Ng, 2 August 2008; others: 1 male molt (animal lost after molting in laboratory, photo voucher) (ca. 12.9 mm carapace width) ( ZRC 2000.1654), Pulau Manukan, Sabah, Malaysia, coll. 22 June 2000.
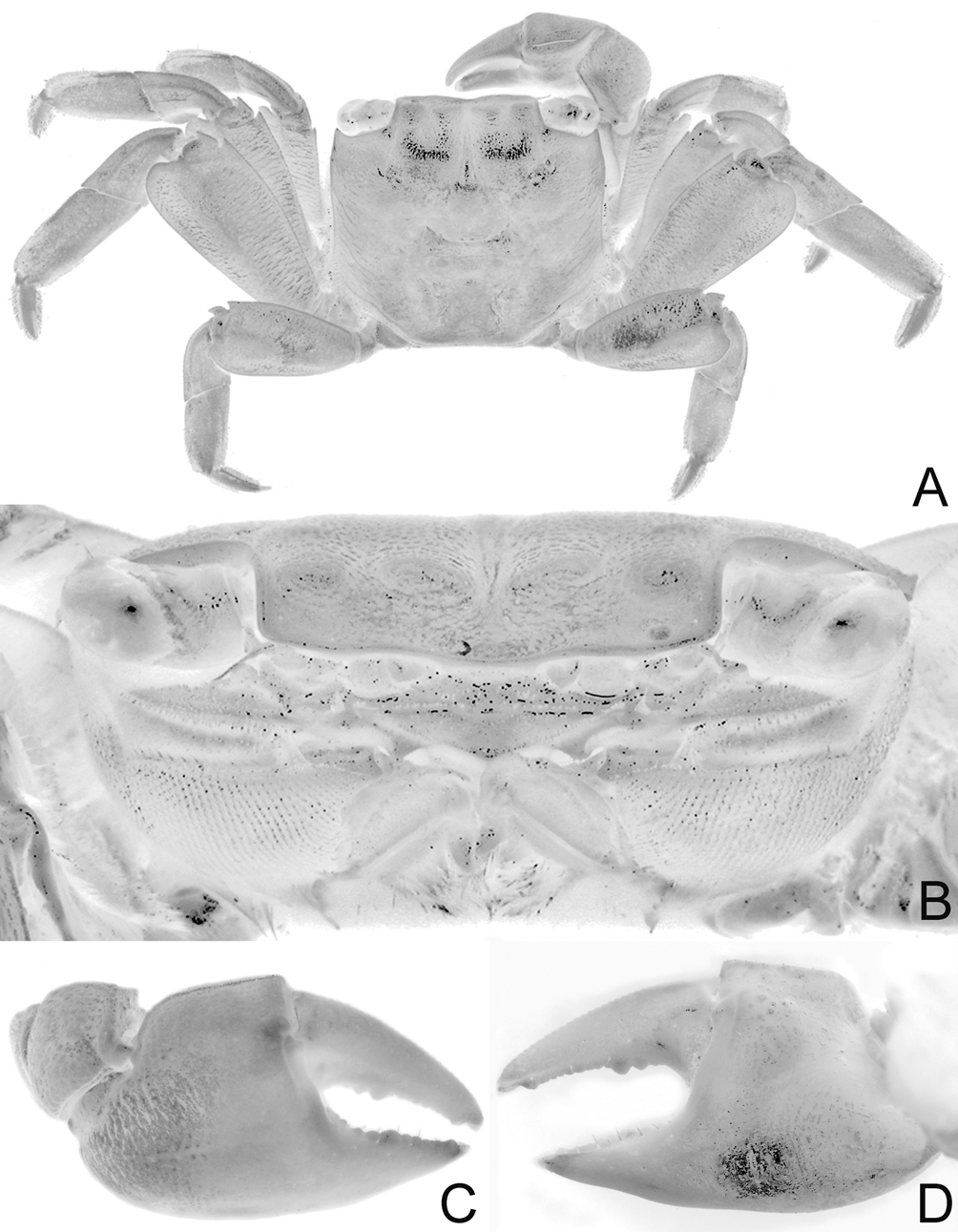
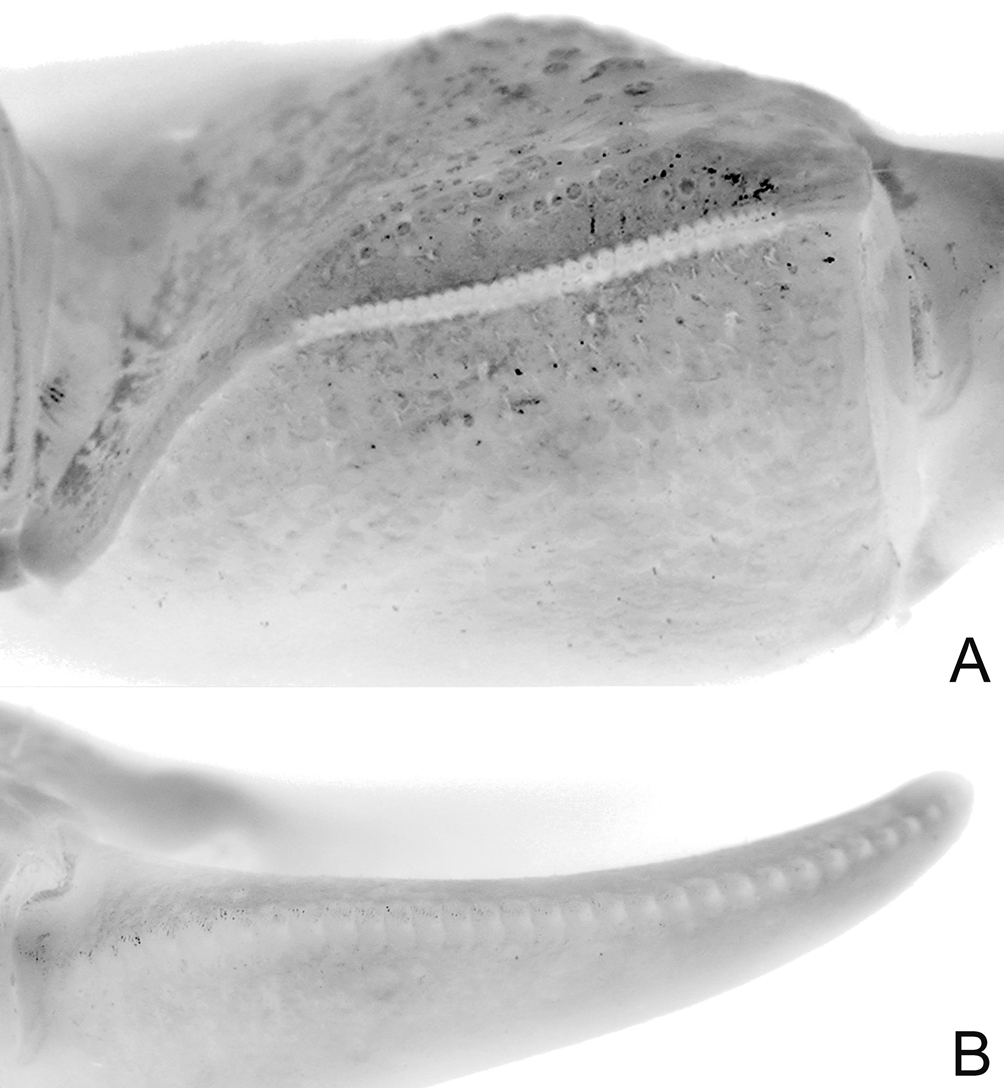
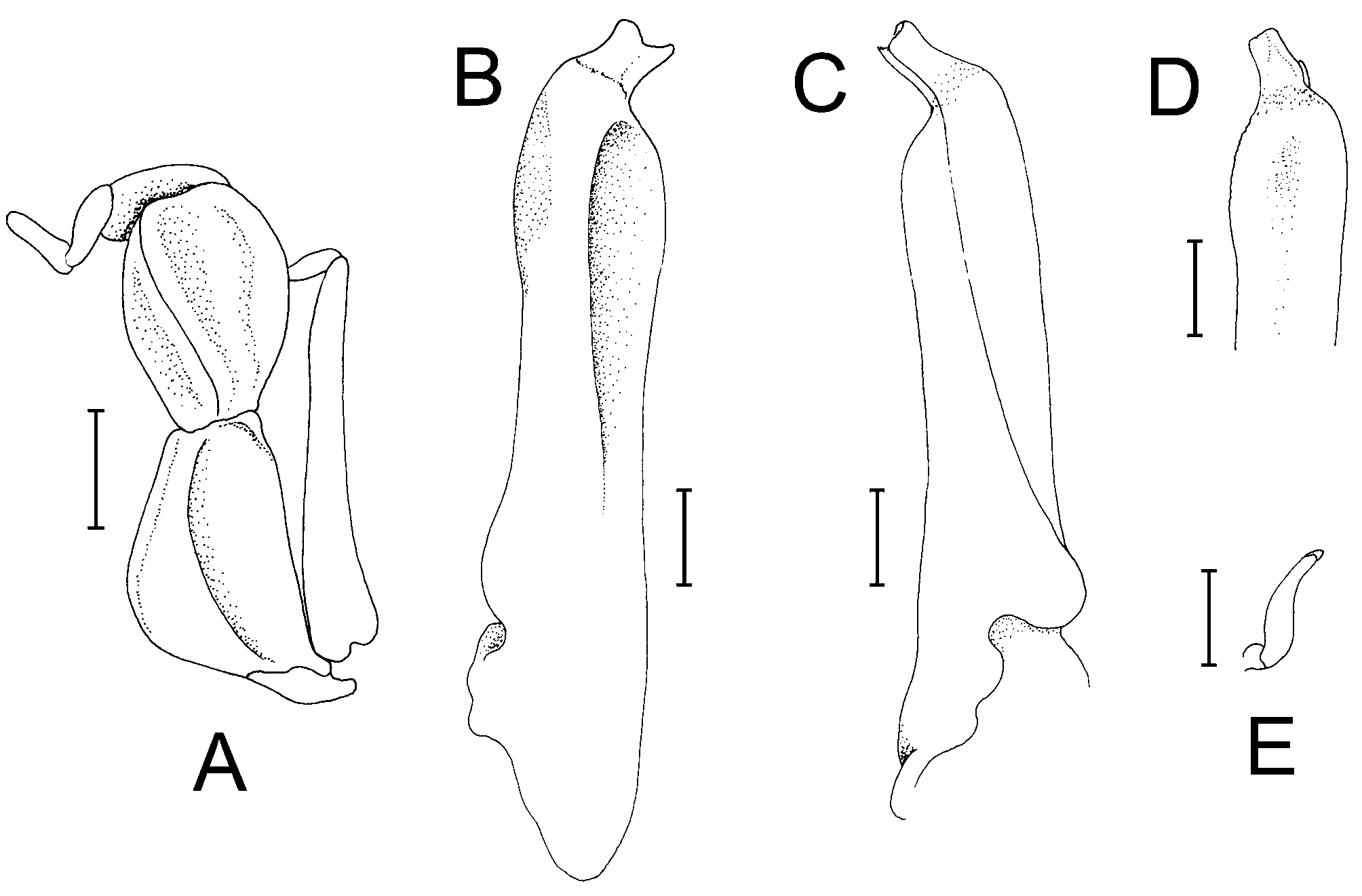
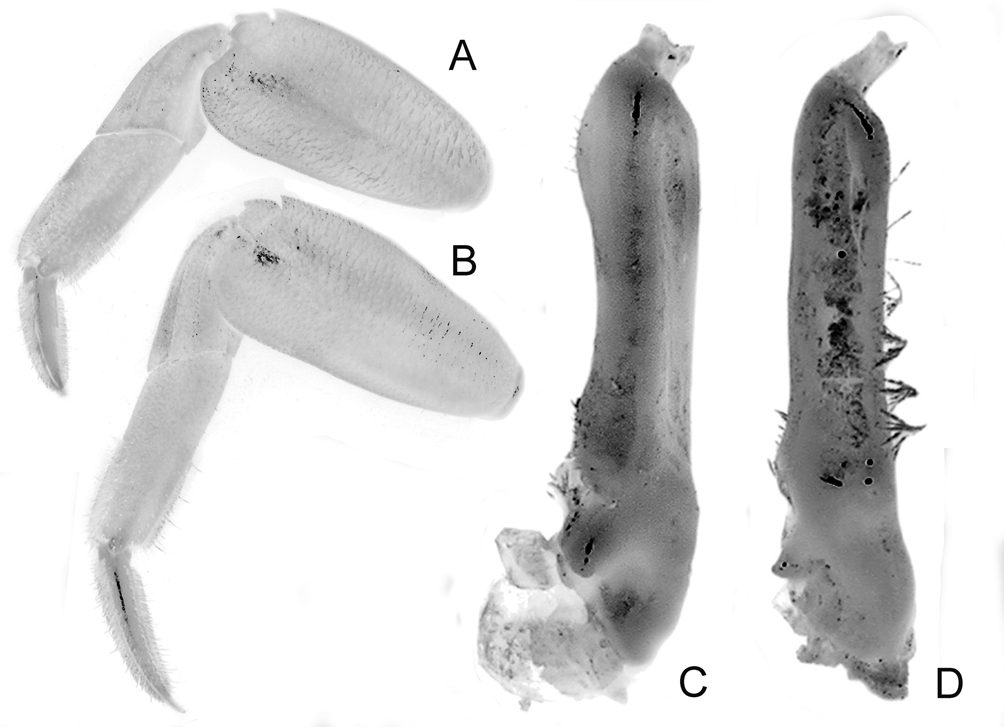
Diagnosis of holotype male. Carapace nearly quadrate, lateral margins diverging slightly toward posterior margin; lateral margins entire, without epibranchial tooth but with shallow concavity where tooth is normally located ( Figs. 2 View FIGURE 2 A, 4A); frontal margin sinuous, weakly 4-lobed ( Figs. 2 View FIGURE 2 A, B, 4A, B); postfrontal crests well developed but not sharp ( Figs. 2 View FIGURE 2 A, B, 4A, B). Outer, inner surfaces of chelae with low, rounded granules or almost smooth, without prominent striae or granules ( Figs. 2 View FIGURE 2 B, 4C, D); dorsal margin of palm with distinct longitudinal crest, proximal third with small, rounded granules, most of distal two-thirds composed of 34 small peg-like, pectinated teeth, distalmost part with 2 or 3 very low, rounded, uneven ridges; inner surface, just behind main crest with about 3 short, oblique subsidiary rows of low tubercles ( Figs. 2 View FIGURE 2 B, 5A); dorsal margin of dactylus with row of 30 tubercles, each tubercle with conical apex surrounded by circular rings, tubercles relatively larger, more closely spaced along proximal part ( Fig. 5 View FIGURE 5 B); cutting edge of dactylus with large submedian tooth followed by 2 smaller proximal ones, distal half with numerous denticles ( Figs. 2 View FIGURE 2 B, 4C, D); cutting edge of pollex with large submedian tooth, smaller ones on both sides ( Figs. 2 View FIGURE 2 B, 4C, D). Ambulatory legs with merus, propodus, dactylus relatively short; margins of dactylus, distal half of propodus lined with dense short, stiff setae ( Figs. 2 View FIGURE 2 A, 3, 4A, 9A). G1 with distal pectinated part relatively long, subtruncate, tip clefted, lower, upper parts subequal ( Figs. 7 View FIGURE 7 B–D, 9C); G2 very short, with cup-like tip ( Fig. 7 View FIGURE 7 E).
Colour. Dorsal surface of carapace purple to brown, with reticulate pattern of green spots and lines; ambulatory legs purple to dark brown with streaks of green; chelae purple with orangish-purple fingers in smaller females; palm of large male green with purplish-orange fingers; eyes iridescent green ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ).
Etymology. The species is named in honor of Prof. Pitiwong Tantichodok from Walailak University, the project leader of the first author. He has devoted his life to teaching marine biology and ecology to students from all over Thailand, especially through his marine ecology summer course, now in its 18th year. He is also an old friend of the second author.
Remarks. Lithoselatium tantichodoki new species, is similar to L. kusu (from Singapore and Malaysia) with regards to its relatively more quadrate carapace and the morphology of the male abdomen and third maxilliped.
However, the proportions of the ambulatory legs of the two species are quite different, with the propodus and dactylus of L. tantichodoki new species ( Figs. 2 View FIGURE 2 A, 4A, 9A) distinctly shorter than those in L. kusu ( Figs. 8 View FIGURE 8 A, 9B) (e.g., length to width ratios of propodus and dactylus of third ambulatory leg of L. tantichodoki is ca. 2.5 and 3.1, respectively, against ratios of ca. 3.1 and 6.0 for L. kusu , respectively). This is especially apparent when specimens of similar sizes are compared. This difference is also valid for the adult and juvenile females of L. tantichodoki new species. A small male (photographed but not preserved) had a similar colour pattern to the juvenile female and the ambulatory legs were slightly longer. Proportions in the moult of a larger male also agree with the type series. The differences between individuals of the same species are therefore not significant. Unfortunately, because only one adult male of L. tantichodoki new species, was obtained, the variation in the structure and number of granules and tubercles on the dorsal margin of the palm and dactylus of the male chela cannot be determined. From what is known for L. pulchrum and L. kusu (see Schubart et al. 2009), this is not likely to be significant.
The G1 structures of L. tantichodoki new species, and L. kusu are different. In L. tantichodoki new species, the G1 is relatively stouter and the margins more sinuous, with the pectinated distal part proportionately shorter ( Figs. 7 View FIGURE 7 B–D, 9C) (relatively more slender, the margins subparallel and almost straight, with the pectinated part proportionately longer in L. kusu , Fig. 9 View FIGURE 9. A, C D; Schubart et al. 2009: fig. 16C–F). The G1 of L. tantichodoki new species, actually resembles more closely that of L. pulchrum from Taiwan ( Schubart et al. 2009: fig. 12D–G). While the number of tubercles on the dactylar finger of the chela is almost the same in L. tantichodoki new species, and L. kusu , there is a difference in their structure and arrangement. In L. tantichodoki new species, the tubercles are relatively more rounded and the spacing between them increases rapidly towards the tip ( Fig. 5 View FIGURE 5 B). In the case of L. kusu , the tubercles are more quadrate, are packed more closely together and the distance between them does not change as rapidly towards the tip ( Fig. 9C View FIGURE 9. A, C ).
The colour patterns of both species are similar ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ; see Schubart et al. 2009: fig. 13 for L. kusu ), with the spots on smaller specimens of L. tantichodoki new species, less well defined and tend to form a reticulate pattern ( Fig. 3 View FIGURE 3 ); while in adults, the pattern becomes less distinct; with the palm of the chela greenish ( Fig. 2 View FIGURE 2 ) (almost completely uniformly purple in L. kusu ; Schubart et al. 2009: fig. 13).
Schubart et al. (2009) noted that one generic feature of Lithoselatium was that a small part of sternite 8 could be observed when the male abdomen is closed. This character is present in L. tantichodoki new species, but the sternite can only be discerned when the setae along the sides of the abdomen are denuded ( Fig. 6 View FIGURE 6 B).
Ecology. Lithoselatium tantichodoki new species, is so far known from Ko Wang Nok and Ko Tan in the Gulf of Thailand ( Fig. 1 View FIGURE 1 ). As in L. kusu , the new species was found under coarse rocks and coral rubble well above the water line in all localities. On Ko Wang Nok, the species share its habitat with Metopograpsus frontalis (Grapsidae) , Leptodius nigromaculatus (Xanthidae) and Epixanthus frontalis (Oziidae) . On Ko Tan, the species is sympatric with Parasesarma pictum (Sesarmidae) , Metopograpsus frontalis (Grapsidae) and Ozius guttatus (Oziidae) .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
Genus |