
Babakina, Roller, 1973
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2007.00331.x |
|
persistent identifier |
https://treatment.plazi.org/id/D00787A3-116D-B270-FC4E-5244FA30A5DE |
|
treatment provided by |
Felipe |
|
scientific name |
Babakina |
| status |
|
PHYLOGENY OF BABAKINA View in CoL
Twenty species of aeolid nudibranchs were considered for the present analyses. All were included in the final analysis. Twenty-seven morphological characters were considered for the present study. Table 2 contains a list of all morphological characters considered. The complete character matrix for species of Babakina is shown in Table 3. Phylogenetic analyses were performed using the program Phylogenetic Analysis Using Parsimony (PAUP) version 4.0b 10 ( Swofford, 2001) using the heuristic algorithm ( TBR branch swapping option), set at maximum parsimony. One thousand replicates were run with starting trees obtained using stepwise addition.
In cases where a taxon had two states for a given character they were treated as uncertain. Both ACCTRAN and the DELTRAN optimizations were used for character transformation. In both cases all characters were treated as unordered with the exception of character 11 (radular teeth number) and were polarized using Notaeolidia gigas Eliot, 1905 as the outgroup species, as members of the Notaeolidiidae are considered to be among the most basal aeolids ( Wägele & Willan, 2000). For a preliminary analysis of the position of the genus Babakina within the Aeolidina, we used several species of aeolids of different families. Aeolidiidae : Aeolidiella alderi (Cocks, 1852) ; Berghia verrucicornis ( Costa, 1864) and Spurilla neapolitana (Delle Chiaje, 1823) . Facelinidae : Caloria elegans (Alder & Hancock, 1845) ; Cratena peregrina (Gmelin, 1791) ; Dicata odhneri (Schmekel, 1967) ; Dondice banyulensis (Portmann & Sandmeier, 1960a) , Facelina auriculata (Müller, 1776) ; Favorinus branchialis (Rathke, 1806) and Pruvotfolia pselliotes (Labbé, 1923) . Flabellinidae : Calmella cavolini (Vérany, 1846) , Flabellina affinis (Gmelin, 1791) ; Flabellina capensis (Thiele, 1925) and Flabellina ischitana (Hirano & Thompson, 1990) . Piseinotecidae : Piseinotecus gaditanus Cervera, García-Gómez & García, 1987 .
Character 16 (presence or absence of a bursa copulatrix) was excluded from the final analysis as its distribution and homology remain poorly understood within the Aeolidina. The bursa is usually the site of exogenous sperm reception following copulation and
1 Notal brim: 0 – present; 1 – interrupted; 2 – absent.
2 Foot corners: 0 – tentacular; 1 – angular; 2 – rounded.
3 Body: 0 – narrow; 1 – wide.
4 Cerata: 0 – arise directly from notum; 1 – arise from elevated peduncles.
5 Ceratal number: 0 – usually less than 100 cerata per side of the body; 1 – numerous, with many more that 100 cerata per side.
6 First ceratal cluster: 0 – rows, present as a series of rows; 1 – arch, forming a horseshoe-shaped arch.
7 Second ceratal cluster: 0 – rows, present as a series of rows; 1 – arch, forming a horseshoe-shaped arch.
8 Anus: 0 – pleuroproctic; 1 – cleioproctic; 2 – acleioproctic.
9 Rhinophoral base: 0 – divided; where both rhinophores have separate point of insertion; 1 – united, where the two rhinophores join and share a common base.
10 Rhinophoral ornamentation: 0 – smooth, with no sign or ornamentation, may occasionally appear wrinkled; 1 – annulate, with a series of well-separated rings; 2 – perfoliate, with a series of crowded lamellae; 3 – papillate with series of elongate papillae; 4 – swelling; with one or more inflated areas along the rhinophoral length.
11 Radula: 0 – multiseriate with several row of lateral teeth; 1 – triseriate with a lateral tooth flanking either side
of the rachidian row; 2 – uniseriate, with only a single rachidian row.
12 Lateral teeth: 0 – denticulate, with a series of denticles along the inner masticatory edge; 1 – smooth, with no denticles along cutting edge.
13 Rachidian tooth shape: 0 – cuspidate, with a series of denticles flanking a triangular cusp; 1 – pectinate, forming a series of comb-like denticles without a larger central cusp.
14 Rachidian tooth: 0 – denticulate; 1 – smooth, without denticles.
15 Radular teeth: 0 – symmetrical, with the same number of denticles on either side of the primary cusp; 1 – asymmetrical, with different numbers of denticles on other side of the cusp.
16 Jaw denticles: 0 – numerous rows, with multiple rows of denticles along the masticatory margin; 1 – single row, with only one row of denticles; 2 – absent, with no denticles along margin.
17 Receptaculum seminis arrangement: 0 – proximal, in closer proximity to the hermaphroditic gland than to the genital aperture; 1 – distal, near the genital aperture.
18 Receptaculum seminis shape: 0 – simple; 1 – bilobed.
19 *Bursa copulatrix: 0 – present; 1 – absent.
20 Bursa copulatrix duct: 0 – straight; 1 – bent.
21 Bursa copulatrix shape: 0 – pyriform; 1 – narrow.
22 Bursa copulatrix size: 0 – large; 1– small.
23 Penis: 0 – unarmed, without cuticular structures; 1 – armed, with cuticular hooks.
24 Penial papilla: 0 – narrow, 1 – conial, 2 – bulbous.
25 Penial glands: 0 – absent, devoid of any auxiliary glandular structures; 1 – present, with secondary glandular structures.
26 Prostate diameter: 0 – wider in some parts; 1 – uniform diameter throughout.
27 Food: 0 – hydroids; 1 – sea anemones; 2 – eggs, with opisthobranch eggs as the primary food source.
may also serve a gametolytic function. The wall of the bursa is lined with columnar endothelial cells. There are both secretory and resorptive cells ( Schmekel, 1971). The bursa is present in some aeolids ( Gosliner, 1994) but this was not confirmed histologically. Schulze & Wägele (1998) give a detailed description of the internal anatomy and the histology of organs and systems of Flabellina affinis . According to these authors, this species has two receptacula, but neither of them shows a gametolytic activity. Some years before, Medina et al. (1988) described the ultrastra- structure of the bursa copulatrix and receptaculum seminis in Hypselodoris midatlantica Gosliner, 1990 (as H. tricolor ). These authors confirmed a gametolytic fuction in the bursa copulatrix, with a single epithelium lining a lumen filled with a heterogeneous material including remnants of degenerative gametes. A thin layer of connective tissue externally surrounds the epithelium. In the luminal material sperm tails, myelinic formations and homogeneously electron-dense bodies are distinguished. In the case of the receptaculum seminis, it is pyriform and the lumen is filled with abundant spermatozoa that are orientated radially. The sperm heads are embedded in the lining epithelium and the tails are directed toward the centre of the lumen. In this chromodoridid species an histological description for these structures exists, but it is necessary to carry out additional studies to clarify possible homologies within Nudibranchia , prior to a phylogenetic analysis.
Morphological data were compiled using MacClade, version 4.08 ( Maddison & Maddison, 2005). Synapomorphies were mapped using the character trace option in MacClade using the majority rule tree from the PAUP analysis. Bremer analyses were performed on the strict consensus tree to estimate branch support ( Bremer, 1994).
CHARACTERS
To determinate the phylogenetic relationships of species of the genus Babakina , 27 characters were examined for 20 taxa. The character states are indicated as follows: 0: the presumed plesiomorphic condition; 1, 2, 3: apomorphic conditions. For character states that are not applicable ‘–’ is used and, for character states that are very variable in a taxa, ‘?’ is used. The character states for the data examined for the present study can be found in Table 2.
1. Notal brim: 0 – present; 1 – interrupted; 2 – absent. The presence of a rim of tissue along the dorsolateral margins of the body has been considered as a plesiomorphic feature within the Aeolidina ( Odhner, 1939). This feature is present in Babakina and N. gigas (0). These taxa are considered very primitive within Aeolidina. Members of the genus Flabellina Voigt, 1834 , Calmella cavolini and Piseinotecus gaditanus have a discontinuous notal brim (1) and they group together in the same clade. In the remaining taxa, no vestige of the notal brim remains (2).
2. Foot corners: 0 – tentacular; 1 – angular; 2 – rounded. The anterior region of the foot in Notaeolidia is rounded (2). In the case of the genera Piseinotecus , Aeolidiella and Cratena , this region is angular (1). In the rest of the species in study, the foot corners are tentacular (0).
3. Body: 0 – narrow; 1 – wide. The species of Aeolidiidae and Notaeolidia possess a wide body (1). The rest of the species in this study possess a narrow body (0).
4. Cerata: 0 – arise directly from notum; 1 – arise from peduncles. In F. ischitana , Calmella cavolini and Piseinotecus gaditanus , the ceratal cluster emerges from stalked clusters, which are well elevated from the notum. The other taxa have the cerata arranged in linear rows. In the case of the Flabellina affinis , there is considerable variability; the arrangement of the cerata can be arranged separately or in rows and/or groups that are either placed on the notal ridge or on peduncles ( Schulze & Wägele, 1998).
5. Ceratal number: 0 – usually less than 100 cerata per side of the body; 1 – numerous, with many more that 100 cerata per side. The cerata of the species of Spurilla , Berghia and Aeolidiella are far more numerous that in the other taxa.
6. First ceratal cluster: 0 – rows, present as a series of rows; 1 – arch, forming a horseshoeshaped arch. In the case of the genera Spurilla , Berghia , Dicata , Cratena, Favourinus and Dondice , the first ceratal cluster forms a horseshoe-shaped arch (1). The remaining species in study have the first ceratal cluster arranged in rows (0).
7. Second ceratal cluster: 0 – rows, present as a series of rows; 1 – arch, forming a horseshoeshaped arch. The vast majority of ingroup taxa that have the arrangement of the first ceratal cluster arranged in an arch, and have the second ceratal cluster in arch as well. Only Cratena has a first ceratal cluster in arch with the second cluster forming a row (0).
8. Anus: 0 – pleuroproctic; 1 – cleioproctic; 2 – acleioproctic. In the species of Notaeolidiidae and the most primitive species of Flabellinidae , the anus is situated in the pleuroproctic position ( Gosliner & Willan, 1991). The species of Flabellina , Calmella , Notaeolidia and Babakina are pleuroproctic (0). Of the taxa included in this analysis only Piseinotecus has the anus in an acleioproctic position (2). The remaining taxa are cleioproctic (1).
9. Rhinophoral base: 0 – divided; where both rhinophores have a separate point of insertion into the dorsal surface of the head; 1 – united, where the two rhinophores join and share a common base. This character is an external feature typical of genus Babakina .
10. Rhinophoral ornamentation: 0 – smooth, with no sign of ornamentation, may occasionally appear wrinkled; 1 – annulate, with a series of well-separated rings; 2 – perfoliate, with a series of crowded lamellae; 3 – papillate with series of elongate papillae; 4 – swelling; with one or more inflated areas along the rhinophoral length. The simple condition, a smooth rhinophore, is considered to represent the ancestral state (0), as it has less sensory surface area. The rhinophores in derived species are generally ornamented. Flabellina ischitana and F. affinis , as well as in Dondice , Pruvotfolia and Facelina , possess annulate rhinophores (1). Spurilla neapolitana and members of the genus Babakina have perfoliate rhinophores (2). Babakina caprinsulensis has incomplete lamellae on its perfoliate rhinophores. Species of Berghia have papillate rhinophores (3), and in the case of Favorinus the rhinophores have a subapical inflated area (4).
11. Radula: 0 – multiseriate with several rows of lateral teeth; 1 – triseriate, with a lateral tooth flanking either side of the rachidian row; 2 – uniseriate, with only a single rachidian row. The multiseriate radula is considered to represent the plesiomorphic state (0); Notaeolidia has a multiseriate radula. The species of Flabellina and Calmella cavolini have a triseriate radula (1). The uniseriate radula is the apomorphic state (2). This character was treated as ordered in the analysis based on the fact that the successive loss of lateral teeth has been well documented in aeolid phylogeny ( Wägele & Willan, 2000).
12. Lateral teeth: 0 – denticulate, with a series of denticles along the inner masticatory edge; 1 – smooth, with no denticles along cutting edge. In most species of Flabellina , the lateral radular teeth bear a series of denticles along their inner edge. The absence of denticles on the lateral teeth is considered to represent a derived feature within Flabellina ( Gosliner & Willan, 1991) . Notaeolidia bears denticles along the inner masticatory edge of the lateral teeth (0). This character is treated as non-applicable in species with uniseriate radula.
13. Rachidian tooth shape: 0 – cuspidate, with a series of denticles flanking a triangular cusp; 1 – pectinate, forming a series of comb-like denticles without a larger central cusp. In species of Spurilla , Berghia and Aeolidiella , the rachidian teeth are pectinate, with denticles on each side of a triangular central cusp.
14. Rachidian tooth: 0 – denticulate, 1 – smooth, without denticles. Only Favorinus has a rachidian tooth with a single denticle, with no denticles on either side of the central denticle. The rest of the species included in the analysis have a denticulate rachidian tooth.
15. Rachidian radular teeth: 0 – symmetrical, with the same number of denticles on either side of the primary cusp; 1 – asymmetrical, with different numbers of denticles on either side of the cusp. In Babakina festiva and B. indopacifica , the number of denticles is not equal on either side of the rachidian tooth.
16. Jaws denticles: 0 – with multiple rows of denticles along the masticatory margin; 1 – single row, with only one row of denticles; 2 – absent, with no denticles along margin. Jaws with several rows of denticles on the masticatory border are considered plesiomorphic (0), whereas those with only one row of denticles (1), or no denticles (2), are apomorphic ( Wägele & Willan, 2000). Spurilla , Berghia , Cratena , Dondice and the clade including Pruvotfolia and Facelina have a single row of denticles in the masticatory border (1). Notaeolidia , Aeolidiella , Dicata and Caloria lack denticles along the margin of the masticatory border (2).
17. Receptaculum seminis arrangement: 0 – proximal, in closer proximity to the hermaphroditic gland than to the genital aperture; 1 – distal, near the genital aperture. A proximal receptaculum seminis is considered plesiomorphic (0). Flabellina affinis is the more derived species of Flabellina in the analysis and it is the only one with a distal receptaculum seminis (1).
18. Receptaculum seminis shape: 0 – simple; 1 – bilobed. A bilobed receptaculum is considered apomorphic (1). Flabellina affinis is the only taxon with a bilobed receptaculum in this study. One lobe is not stalked and another one is smaller and stalked.
19. Bursa copulatrix: 0 – present; 1 – absent. The presence of a proximal receptaculum seminis and a distal bursa copulatrix is considered a plesiomorphic state ( Ghiselin, 1966; Gosliner, 1981; Schmekel, 1985; Mikkelsen, 1996). Notaeolidia , Flabellina capensis , F. ischitana , Dicata and the species of Babakina have a bursa copulatrix, and are considered plesiomorphic (0). The remaining species included in the analysis lack a bursa (1). This character was deleted from the final analysis owing to the fact that the bursa has often been overlooked and homology of this structure remains open to some question.
20. Bursa copulatrix duct: 0 – straight; 1 – bent. In two species, Flabellina capensis and Babakina caprinsulensis , the bursa has a sharp bend. This is considered plesiomorphic (0). In remaining species the duct is simply curved or straight (1). In the species with bursa copulatrix absent, this character is treated as nonapplicable.
21. Bursa copulatrix shape: 0 – pyriform; 1 – narrow. In Babakina festiva and B. indopacifica , the bursa copulatrix has a narrow shape (1). In the remaining species with bursa copulatrix, it has a pyriform shape (0). In those species with bursa copulatrix absent, this character is treated as non-applicable.
22. Bursa copulatrix size: 0 – large; 1 – small. Presence of a small bursa is a feature typical of B. indopacifica , where the bursa copulatrix is reduced (1). In the species lacking a bursa copulatrix, this character is treated as non-applicable.
23. Penis: 0 – unarmed, without cuticular structures; 1 – armed, with cuticular hooks. The absence of spines within the vas deferens is considered the plesiomorphic state (0) ( Gosliner, 1994; Wägele & Willan, 2000). Pruvotfolia pselliotes and Facelina auriculata ( Alder & Hancock, 1855) have a penis with cuticular spines (1). The rest of the species in the study have an unarmed penis.
24. Penial papilla: 0 – narrow; 1 – conical; 2 – bulbous. The species Berghia verrucicornis , the clade including Flabellina ischitana , F. affinis , Calmella and Piseinotecus , as well as the clades including Babakina festiva have a narrow penial papilla (0). Aeolidiella , Dicata, Pruvotfotlia , Dondice , Babakina caprinsulensis and B. anadoni have a conical penial papilla (1). The remaining species in this study present a bulbous penial papilla (2).
25. Penial glands: 0 – absent, devoid of any auxiliary glandular structures; 1 – present, with secondary glandular structures. A simple penis, without penial glands is considered plesiomorphic (0). Facelina auriculata is the only taxon in this analysis that has penial glands (1).
26. Prostate diameter: 0 – wider in some parts; 1 – uniform diameter throughout. Notaeolidia , Cratena , Favorinus , Dondice and the species of Babakina have a uniform diameter of the prostate (1); the other species have a welldifferentiated prostatic section divided into several parts (0)
27. Food: 0 – hydroids; 1 – sea anemones; 2 – eggs. The vast majority of species included in the analysis feed on hydroids (0). Spurilla , Berghia and Aeolidiella feed on sea anemones (1) while Favorinus feeds on opisthobranch eggs (2).
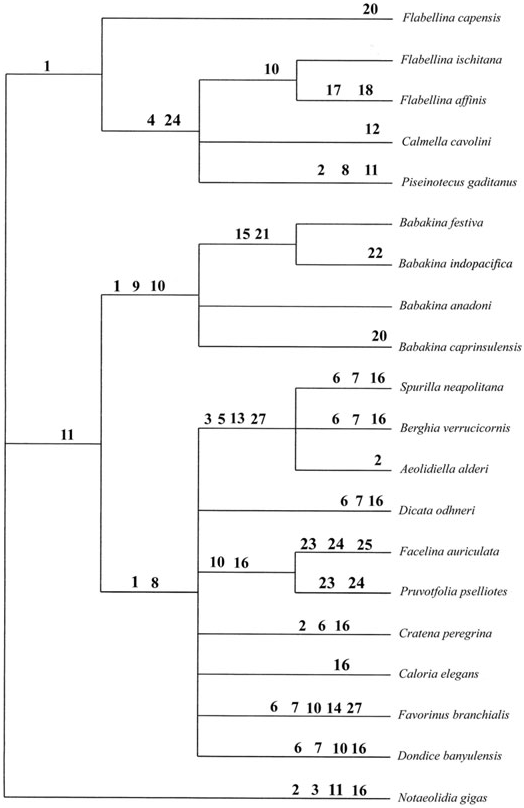
In the analysis presented ( Fig. 6 View Figure 6 ), The heuristic search of the data matrix set produced 16 most parsimonious trees, obtained with 58 steps, and had a consistency index (CI) of 0.63, a retention index (RI) of 0.75 and a homoplasy index (HI) of 0.36.
The resulting phylogeny shows phylogenetic relationships that are rather weakly supported by Bremer values of 1 or 2 ( Fig. 7). Babakina represents a clade that is moderately well supported by two synapomorphies: united rhinophores (9) and perfoliate rhinophores (10) and has a Bremer support value of 2. The clade including B. festiva and B. indopacifica is supported by two synapomorphies: different numbers of denticles on other side of the cusp of the rachidian tooth (15) and bursa copulatrix with narrow shape (21). Additionally, B. indopacifica has an auto- pomorphic feature: a reduced bursa copulatrix that is characteristic of this species (22). Babakina caprinsulensis has one autopomorphy: a bent bursa duct (20). Within Babakina , B. anadoni and B. caprinsulensis form a trichotomy with the clade containing B. festiva and B. indopacifica . The sister group relationship of B. festiva and B. indopacifica has a Bremer support value of 2.
Babakina is the sister group to Aeolidiidae plus Facelinidae . Aeolidiidae is a monophyletic group and is weakly supported as a clade (Bremer support value of 2). Aeolidiidae is supported by four synopomophies: a broad body (3), a high number of cerata (5), a pectinate rachidian tooth (13) and feeding on sea anemones (27). However, Facelinidae is not supported as a clade in the present analysis. Relationships within Facelinidae are not supported, most probably due to incomplete taxon sampling.
The Babakina / Aeolidiidae / Facelinidae clade is supported by one synapomophy: uniseriate radula (11), and has a Bremer support value of 1. The clade including Aeolidiidae and Facelinidae is supported by a Bremer value of 1 and two synapomorphies: absence of a notal brim (1) and a cleioproctic anus (8).
Flabellinidae is weakly supported (Bremer support value of 1), but monophyletic when Calmella and Piseinotecus are included as members of this taxon. They are supported by the presence of a single synapomorphy: presence of an interrupted notal brim (1). The clade including Flabellina ischitana , F. affinis , Calmella and Piseinotecus is supported by a Bremer value of 1 and two synapomorphies: cerata arising from peduncles (4) and a narrow penial papilla (24).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.