
Urticina, Ehrenberg, 1834
|
publication ID |
https://doi.org/10.1080/00222930600703532 |
|
persistent identifier |
https://treatment.plazi.org/id/CF17AF78-FFB5-386F-FDFA-B775FBAFCDE9 |
|
treatment provided by |
Felipe |
|
scientific name |
Urticina |
| status |
|
Genus Urticina View in CoL
Diagnosis (modified from Carlgren 1949)
Actiniidae with well-developed pedal disc. Column with adhesive or non-adhesive verrucae or without these. Fosse well developed. Sphincter strong, circumscribed. Tentacles short, stout, their longitudinal muscles ectodermal to mesogloeal. Radial muscles of oral disc ectodermal to mesogloeal. Numerous perfect mesenteries arranged as a rule decamerously or more irregularly 11-, 12-, 13-, or 14-merously. Usually 10–20 oldest pairs sterile, rarely only six pairs. Basitrichs of the actinopharynx much larger than those of the tentacles and size ranges do not overlap. Same number of mesenteries proximally and distally. Cnidom: spirocysts, basitrichs, microbasic p-mastigophores A, and microbasic p-mastigophores B.
Urticina crassicornis ( Müller, 1776) View in CoL
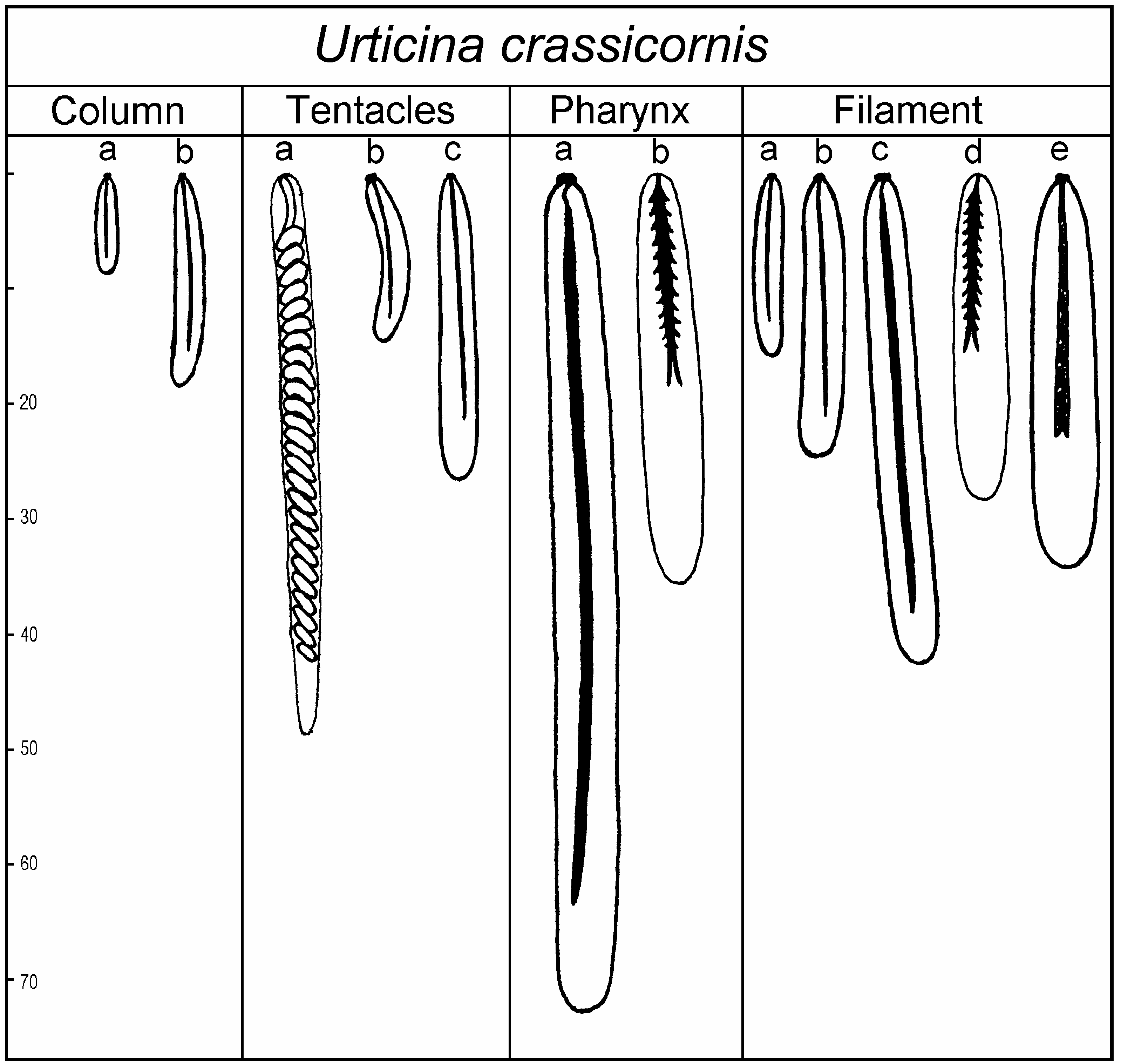
( Figures 6 View Figure 6 , 15A, B View Figure 15 )
Actinia crassicornis Müller 1776, p 231 View in CoL .
Urticina felina: McMurrich 1911, p 65 View in CoL .
Urticina felina crassicornis: Carlgren 1921, p 170 View in CoL (synonymy).
Tealia felina var. crassicornis: Stephenson 1935, p 150 View in CoL .
? Tealia crassicornis: Hand 1955, p 72 View in CoL (part).
Not Tealia crassicornis: Chia and Spaulding 1972, p 206 View in CoL ; Sebens and Laakso 1977, p 165; Widersten 1976, p 865.
Material examined
East Kamchatka: Avacha Bay, Starichkov Island , 14 m, 21 September 2004, one specimen ( KBPIG 271 /12); Kekkurny Point, 12 m, 14 July 2003, three specimens ( KBPIG 274 /18); Bezymyannaya Bay , 9–12 m, 1 October 2004, three specimens ( KBPIG 272 /16, 273/17); November 2004, one specimen ( KBPIG 275 /19). Commander Islands , Medny Island : Drovenskoy Point, intertidal pool, 4 August 1995, three specimens (5/1, 62/8) ; 5 m, 23 July 1995, two specimens ( KBPIG 58 /5); 15 m, one specimen ( KBPIG 61 /7); Glinka Bay, 6 m, 8 August 1995, six specimens ( KBPIG 8 /2, 56/3, 57/4, 60/6); 5 m, 1 August 1995, one specimen ( KBPIG 64 /10); Sivuchi Kamen Kliff, 10 m, 23 July 1992, one specimen ( KBPIG 65 /11); Gladky Point, 5 m, 21 June 1992, one specimen ( KBPIG 163 /13); Gavrilovskaya Bay , 15 m, 9 July 1992, one specimen ( KBPIG 165 /14); Matvea Point, 5– 7 m, 5 July 1992, two specimens ( KBPIG 168 /15) .
Description
External appearance. The typical living specimens are up to 10 cm high and with a tentacular crown up to 10 cm diameter and the column about 6–7 cm diameter. The largest formalin-preserved specimen is 4 cm in diameter and 3 cm high. The circular base is wider than the column and strongly adhesive. The colour is variable, although most specimens range from plain reddish brown to pale orange. Some specimens from the Commander Islands are deep red with irregular longitudinal green patches or strips. The column is smooth, without traces of verrucae or vesicles, and is always clear, without attached sand or other foreign particles. The margin is smooth. There is a deep fosse and a short capitulum.
The oral disk is flat and circular, much paler than the column, usually yellowish or whitish. Short thin red radial bands outline the tentacle bases; the region around the mouth may be reddish, and otherwise the colour of the oral disk is plain.
The tentacles, arranged decamerously up to five cycles, number from 74 in the smallest specimen to 163 in large specimens. Conical tentacles, of the same colour as the disk, are plain-coloured, without bands or other markings. In preserved specimens the tentacles are longitudinally folded, cylindrical, up to 2 mm in diameter and up to 7 mm long. Some may be bifurcated, or even have up to four tips.
Internal structure. A strong circumscribed endodermal sphincter with one central lamella is typical for the genus. Radial muscles of the oral disk and longitudinal muscles of the tentacles are mesogloeal, sometimes ectomesogloeal, and strong. The mesenteries are arranged decamerously and regularly in four cycles, 10+10+20+40, although the last cycle may be incomplete in smaller specimens. Two specimens ( KBPIG 274/18, 275/19) have, in addition, several mesenterial pairs of almost the same size as mesenteries of the fourth cycle. The mesenteries of the first, second and some of the third cycle are perfect. In most specimens only third and fourth cycles of the mesenteries are fertile, although the gonads may be present in the mesenteries of the second cycle in small specimens. The retractor muscles on the oldest cycles are long, weak and diffuse, but restricted on the fourth cycle. The parietobasilar muscles are well developed and form a clear pennon.
The sexes are separate. Embryos were not found in any specimen.
Size and distribution of cnidae (letters in brackets refer to Figure 6 View Figure 6 , all measurements in Mm; N is the proportion of examined specimens that had a particular type of cnida; distribution of all cnidae excepting the smallest basitrichs in column is confirmed on sections):
Column basitrichs (a): 7–10×1.5–2 (very rare), N54/13
basitrichs (b): 14–23×2–3 (common), N513/13
Tentacles spirocysts (a): 18–55×2–3.5 (numerous), N511/11 basitrichs (b): 9–17×1.5–2.5 (common, mostly crescent-shaped), N510/11 basitrichs (c): 17–31×2–3 (common), N511/11
Actinopharynx basitrichs (a): 54–86×5–7 (numerous), N513/13 p-mastigophores A (b): 24–40×5–6 (rare), N512/13
Filaments basitrichs (a): 12–17×1.5–2.5 (numerous), N59/10 basitrichs (b): 15–25×2.5–3.5 (common), N58/10 basitrichs (c): 40–66×4–6.5 (very rare), N58/10 p-mastigophores A (d): 20–30×4–5.5 (numerous), N510/10 p-mastigophores B (e): 28–40×5–6.5 (numerous), N510/10
Habitat
The specimens were recorded from the intertidal zone to 20 m depth, attached to stones or rocks, never buried in sand.
Remarks
Carlgren (1921) examined numerous specimens of this species from many localities in the Arctic seas, and distinguished it by the complete absence of any traces of verrucae on the column. In addition to the northern specimens Carlgren (1921, 1934) assigned several specimens from the north Pacific (the Bering Sea and Bering Island) to this species. Hand (1955) described Tealia crassicornis from California. This is the only record of the species from the Pacific published after Carlgren’s works provided with a description and, in general, corresponding to the species. Hand (1955), however, reported that the Californian specimens occasionally have weakly adhesive verrucae to which small particles of gravel may adhere. This feature never occurs in U. crassicornis (confirmed in the present study by examining many living and preserved specimens). This, associated with its southern location, makes the record from California doubtful. More recent records are confused and deal with one or more other distinct species. For instance, Widersten (1976) described smooth and verrucose specimens, some with 48 marginal verrucae (which never occur in U. crassicornis ), and some with only 68 mesenteries (too few for U. crassicornis ). The species discussed as T. crassicornis by Chia and Spaulding (1972) has non-adhesive vesicles on column and parapet and differs in this feature from U. crassicornis and, probably, from the California species described by Hand (1955).
Although the present specimens are morphologically identical to the northern specimens of U. crassicornis , and have similar nematocysts to those reported by Carlgren (1921), there is still some doubt about the north Pacific specimens belonging to U. crassicornis . Although northern specimens are viviparous ( Carlgren 1921; Stephenson 1935), embryos have not been found in the present specimens. Chia and Spaulding (1972) reported U. crassicornis being oviparous in the northeastern Pacific ( San Juan Island). The species they studied, however, is not conspecific with U. crassicornis ; it is similar to and probably conspecific with U. grebelnyi sp. nov. Chia and Spaulding (1972: 206) compared their findings with the data of Appellöf (1900) ‘‘who reported that in Europe this species releases its gametes freely into the sea’’. However, the species studied by Appellöf (1900) is not conspecific with U. crassicornis and was synonymised with T. felina lofotensis (5 U. eques ) by Carlgren (1921).
Urticina eques ( Gosse, 1860) View in CoL
( Figures 7 View Figure 7 , 15C, D View Figure 15 )
Bolocera eques Gosse 1860, p 351 View in CoL .
Madoniactis lofotensis Danielssen 1890, p 47 View in CoL (part).
Tealia lofotensis: Carlgren 1902, p 42 View in CoL .
Urticina felina lofotensis: Carlgren 1921, p 168 View in CoL (synonymy).
Tealia felina var. lofotensis: Stephenson 1935, p 147 View in CoL .
Urticina eques: Manuel 1981, p 109 View in CoL .
Not Tealia lofotensis: Hand 1955, p 80 View in CoL ; Sebens and Laakso 1977, p 162 (5 Cribrinopsis albopunctata View in CoL sp. nov.).
Material examined
Lofoten , Norway, 12 specimens, coll. Dr D. Schories ( KBPIG 256 /1, 257/2) .
This species has not been recorded from the Pacific. Specimens collected in Lofoten, Norway ( type locality of Madoniactis lofotensis ) are briefly described for comparison with the Pacific species of the genus .
Description
External appearance. Available formalin-preserved specimens are dome-shaped, strongly contracted, with completely retracted tentacles, 28–46 mm diameter and 17–23 mm high. On underwater photos the same specimens have whitish or yellowish ground colour with wide or narrow, irregular, mainly longitudinal red patches. In some specimens the prevailing colour is red. Small non-adhesive verrucae are distinctly visible on the living specimens, especially on the contracted anemones ( Figure 15C View Figure 15 ). The verrucae are of the same colour as the column or paler, and they probably do not become as inflated and blister-like as in U. grebelnyi sp. nov. In preserved specimens the verrucae are difficult to recognize. The oral disk and the tentacles are yellow-whitish, transparent, with the red longitudinal bands outlining tentacle bases on the disk; the tentacles are encircled with the wide white and red bands in the middle. Tentacles are arranged decamerously in five cycles.
Internal structure. The sphincter is strong, circumscribed and circular in cross-section. Radial muscles of the oral disk and longitudinal muscles of the tentacles are mesogloeal. Mesenteries are arranged decamerously, 10+10+20+40, although in the smaller specimens the last cycle may be incomplete. The largest specimen has the following arrangement of the mesenteries: 12+10+22+43+1. Mesenteries from the first to the third cycles may be perfect. The mesenteries of the third and fourth cycles are fertile; in one small specimen a gonad is present also on one mesentery of the second cycle. The retractor muscles are long and diffuse.
The sexes are separate. No embryos were found in any specimen.
Size and distribution of cnidae (letters in brackets refer to Figure 7 View Figure 7 , all measurements in Mm; N is the proportion of examined specimens that had a particular type of cnida; distribution of all cnidae is confirmed on sections):
Column basitrichs (a): 8–11×1.5–2 (rare), N54/4 basitrichs (b): 17–25×2–3 (common), N54/4
Tentacles spirocysts (a): 20–60×2–4 (numerous), N53/3 basitrichs (b): 9–12×1.5–2 (rare), N52/3 basitrichs (c): 24–30×2–3 (numerous), N53/3
Actinopharynx basitrichs (a): 63–95×5–6.5 (numerous), N54/4 p-mastigophores A (b): 22–28×4.5–5.5 (very rare), N52/4
Filaments basitrichs (a): 10–14×2 (common), N54/4 basitrichs (b): 15–24×2–3.5 (common), N54/4 basitrichs (c): 54–80×5.5–7 (very rare), N54/4
p-mastigophores A (d): 22–28×4.5–5.5 (common), N54/4
p-mastigophores B (e): 20–39×4.5–7 (common), N54/4
Remarks
For a long time this northern species was attributed to Madoniactis lofotensis Danielssen, 1890 (recorded under the generic names Urticina or Tealia , or as a variety or subspecies of U. felina ). The possible conspecificity of Danielssen’s species with Bolocera eques Gosse, 1860 was first suggested by Carlgren (1921) who questionably synonymised these species, but used U. felina lofotensis as a valid name. Stephenson (1935, p 143) had no doubt that B. eques is conspecific, but also used Danielssen’s name (as T. felina var. lofotensis ). Finally, Manuel (1981) formally synonymised M. lofotensis with Urticina eques and in the present paper we fully support his conclusion.
Manuel (1981, p 110) supposed that T. lofotensis described by Hand (1955) from California may be not conspecific with the present species: ‘‘Although this anemone [ T. lofotensis: Hand, 1955 ] bears strong external resemblance to the present species, measurement of its nematocysts do not wholly agree’’. Hartog (1986, p 87) found this to be an ‘‘understatement’’. Based on his own (unpublished) studies of the northern species he considered the Californian species distinct and indicated that it requires a new name. Actually, the Pacific species widely known as U. lofotensis and described in detail by Hand (1955) and Sebens and Laakso (1977) does not resemble the European species externally. The Pacific species has white, always well-visible verrucae on the usually uniformly coloured crimson column. In European specimens the verrucae are smaller, usually inconspicuous, they may be whitish, but often of the same colour as the column and the species look completely different ( Figure 15C, D View Figure 15 ). Further, the nematocyst data clearly distinguish the two species: the lectotype of Madoniactis lofotensis (see Carlgren 1921), as well as the present specimens of U. eques have cnidom typical for Urticina , while the cnidom of Pacific specimens is typical for Cribrinopsis ( Figure 11 View Figure 11 ).
Urticina felina ( Linnaeus, 1761) View in CoL
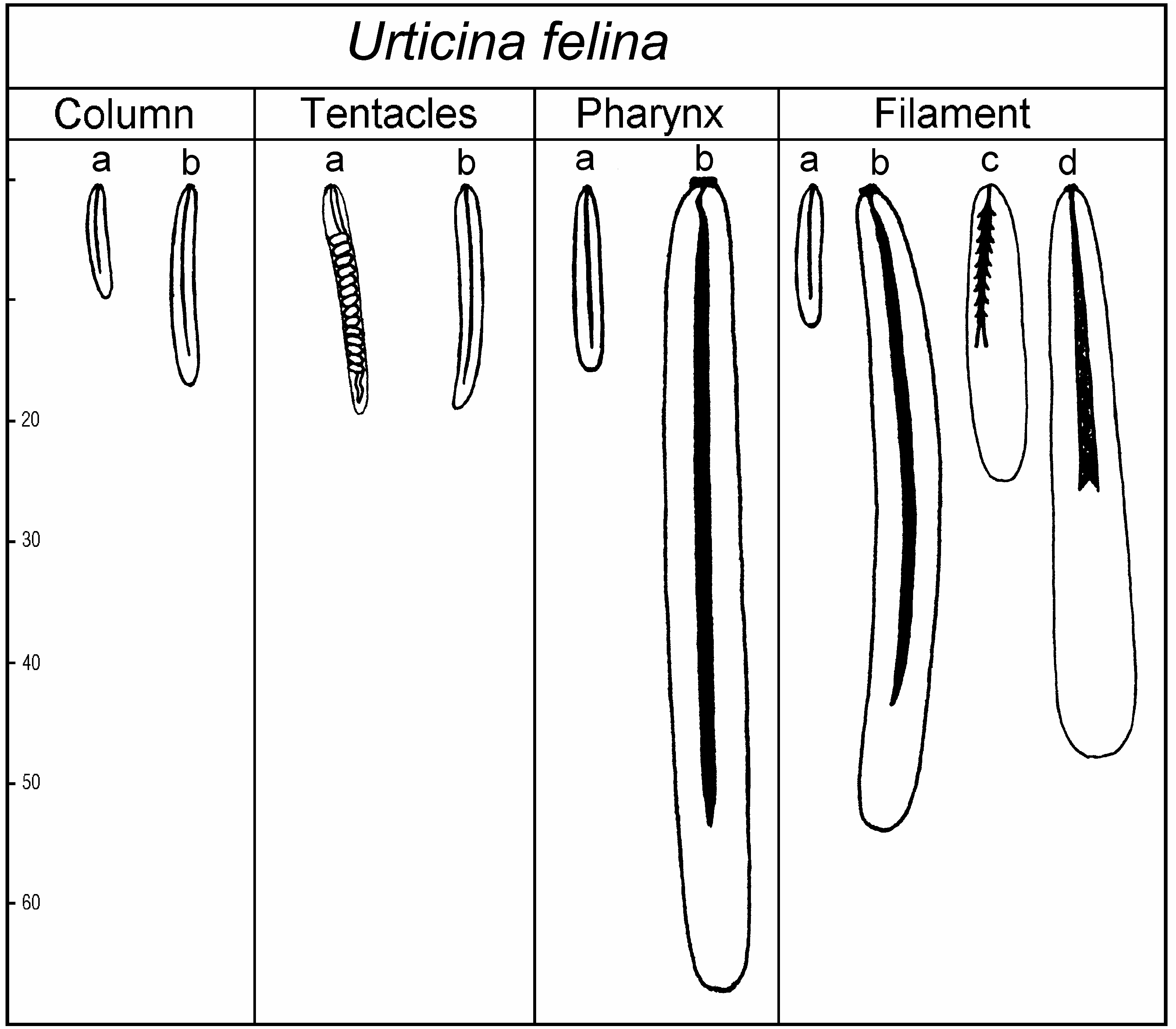
( Figure 8 View Figure 8 )
Priapus felinus Linnaeus 1761, p 510 View in CoL .
Actinia coriacea Cuvier 1798, p 653 View in CoL .
Actinia tuberculata Cocks 1850, p 94 View in CoL .
Urticina felina coriacea View in CoL and tuberculata: Carlgren 1921, p 163 View in CoL (synonymy).
Tealia felina var. coriacea: Stephenson 1935, p 140 View in CoL .
Urticina felina: Manuel 1981, p 106 View in CoL .
Not Tealia coriacea: Hand 1955, p 76 View in CoL .
Material examined
Great Britain, Swansea, the Mumbles Point , intertidal zone, 18 June 1993, three specimens, coll. Dr A. Rzhavsky ( KBPIG 69 /1) .
This species is not present in our material from the northwestern Pacific. We give brief morphological information and nematocysts data of the examined specimens from the British Isles for comparison with other Urticina species.
Description
Preserved specimens, 27–49 mm in diameter and 13–32 mm high, are highly verrucose, covered by gravel and shell particles. They are in agreement with the previous description of this species from northern waters (see synonymy). Mesenteries are arranged decamerously, in four cycles, the last cycle incomplete. The gonads are on the mesenteries from the second to fourth cycles. In one specimen a gonad is present also on one mesentery of the first cycle. The longitudinal muscles of the tentacles and oral disk are from mesoectodermal to mesogloeal.
Size and distribution of cnidae (letters in brackets refer to Figure 8 View Figure 8 , all measurements in Mm; N is the proportion of examined specimens that had a particular type of cnida; distribution of all cnidae is confirmed on sections):
Column basitrichs (a): 7–12×1.5–2 (very rare), N53/3
basitrichs (b): 15–19×2–2.5 (common), N53/3
Tentacles spirocysts (a): 16–32×2–2.5 (numerous), N52/2 basitrichs (b): 16–22×2–2.5 (common), N52/2
Actinopharynx basitrichs (a): 13–18×2–2.5 (very rare), N52/3 basitrichs (b): 54–77×5–6.5 (common), N53/3
Filaments basitrichs (a): 11–13×2 (common), N53/3 basitrichs (b): 55–61×5.5–6 (very rare), N51/3 p-mastigophores A (c): 20–27×3–5 (common), N53/3 p-mastigophores B (d): 29–52×5–7.5 (common), N53/3
Remarks
This is the best defined and well-known species. Although in one specimen we detected a gonad in one mesentery of the first cycle, a feature previously considered as not occurring in Urticina , there is no doubt about its identity. The specimens agree with all descriptions of this species from Europe and have been collected on British shores, where the species is very common ( Stephenson 1935). We agree with Hartog (1986) who had no doubt that Californian Tealia coriacea: Hand, 1955 is a distinct species and requires a new name. Not one of the recorded species in the studied area in the north-west Pacific is similar to U. felina .
Urticina grebelnyi sp. nov.
( Figures 9 View Figure 9 , 10 View Figure 10 , 14 View Figure 14 )
Tealia crassicornis: Chia and Spaulding 1972, p 206 View in CoL ; Sebens and Laakso 1977, p 165.
Not Actinia crassicornis Müller 1776, p 231 View in CoL .
Urticina tuberculata: Zamponi and Acuna 1996, p 3 View in CoL .
Not Actinia tuberculata Cocks 1850, p 94 View in CoL .
Material examined
Holotype: East Kamchatka: Avacha Bay, Starichkov Island , 7.5 m, 25 July 2002 ( KBPIG 223 /6) . Paratypes: East Kamchatka: Avacha Bay, Bezymyannaya Bay , 10 m, 22 August 2000, one specimen ( KBPIG 193 /3); Kekkurny Point , 12 m, 14 July 2003, one specimen ( KBPIG 259 /13); Zhirovaya Bay , 12–13 m, 15 July 2003, one specimen ( KBPIG 260 /14); Starichkov Island , 10 m, 15 September 2002, one specimen ( KBPIG 229 /7); 20 m, 9 September 2003, one specimen ( KBPIG 238 /10). Kronotsky Bay , Morzhovaya Bay , 12 m, 18 May 2003, two specimens ( KBPIG 231 /8) .
Other specimens. East Kamchatka: Avacha Bay, Kazak Point , 6 m, 25 July 2000, one specimen ( KBPIG 191 /1); 3 m, 10 July 2002, two specimens ( KBPIG 214 /4); 10 August 1983, one specimen ( KBPIG 258 /11); Bezymyannaya Bay , 4 m, 22 August 2000, two specimens ( KBPIG 192 /2); Starichkov Island , 10 m, 26 June 2002, one specimen ( KBPIG 215 /5). Kronotsky Bay , Morzhovaya Bay , 8.5 m, 18 May 2003, one specimen ( KBPIG 235 /9). Commander Islands : Bering Island , Podutesnaya Bay , 20 m, 22 July 1991, two specimens ( KBPIG 67 /12); Tundrovaya Bay , 20 m, 18 August 1986, one specimen ( KBPIG 68 /15) .
Description
External appearance. The typical living specimens are large, about 20 cm high, with the cylindrical column about 15 cm diameter, and the extended crown of the tentacles up to 25 cm diameter. Large formalin-preserved specimens are up to 15 cm diameter and up to 10 cm high. In living specimens the strongly adhesive circular pedal disk is always wider than the column, but in the preserved specimens the pedal disk contracts and often becomes narrower than the column. The colour pattern is almost constant and shows only a little variation. The column of all specimens is covered by large irregular patches of green and red, the size of these patches varying; in some specimens the prevailing colour is red, in others green. The pedal disk is the same colour as the column. The whole column is covered by numerous non-adhesive verrucae ( sensu Hartog 1987). In fully expanded living specimens verrucae do not protrude over the surface of the column, and since they often are not marked by their colour, they may be inconspicuous. When the specimen contracts verrucae become large, inflated and thin-walled ( Figure 14C, F View Figure 14 ). Strongly inflated verrucae may be lobed, especially on the margin. The verrucae on the margin are somewhat larger than the columnar verrucae and have white markings on the tops; each marginal verruca corresponds to one exo- or endocoel. Verrucae may be arranged into longitudinal or transverse rows depending on the contraction of the specimen. Marginal and columnar verrucae have the same nematocysts as the rest of the column. There is a deep fosse, up to 1.5 cm, and a capitulum the same height. The fosse has a bright red band running along the parapet, 5–7 mm wide, becoming green toward the bottom. The capitulum is the same colour as the disk.
The oral disk is flat and circular, pale yellow-green, pale lilac, or brownish. Short thin red radial bands outline the tentacle bases; otherwise the colour of the oral disk is plain.
Tentacles are arranged 12-merously or irregularly in five cycles, up to 200 tentacles in large specimens. Usually they are shorter than the radius of the oral disk (up to 4–5 cm long), up to 1 cm diameter near the base, cone-shaped, longitudinally folded in contraction. Rarely some of the tentacles may have bifurcated tips. The tentacles are the same colour as the disk or paler, and are always encircled by a wide lilac, reddish, or brownish band in the middle. Tentacle tips are often the same colour.
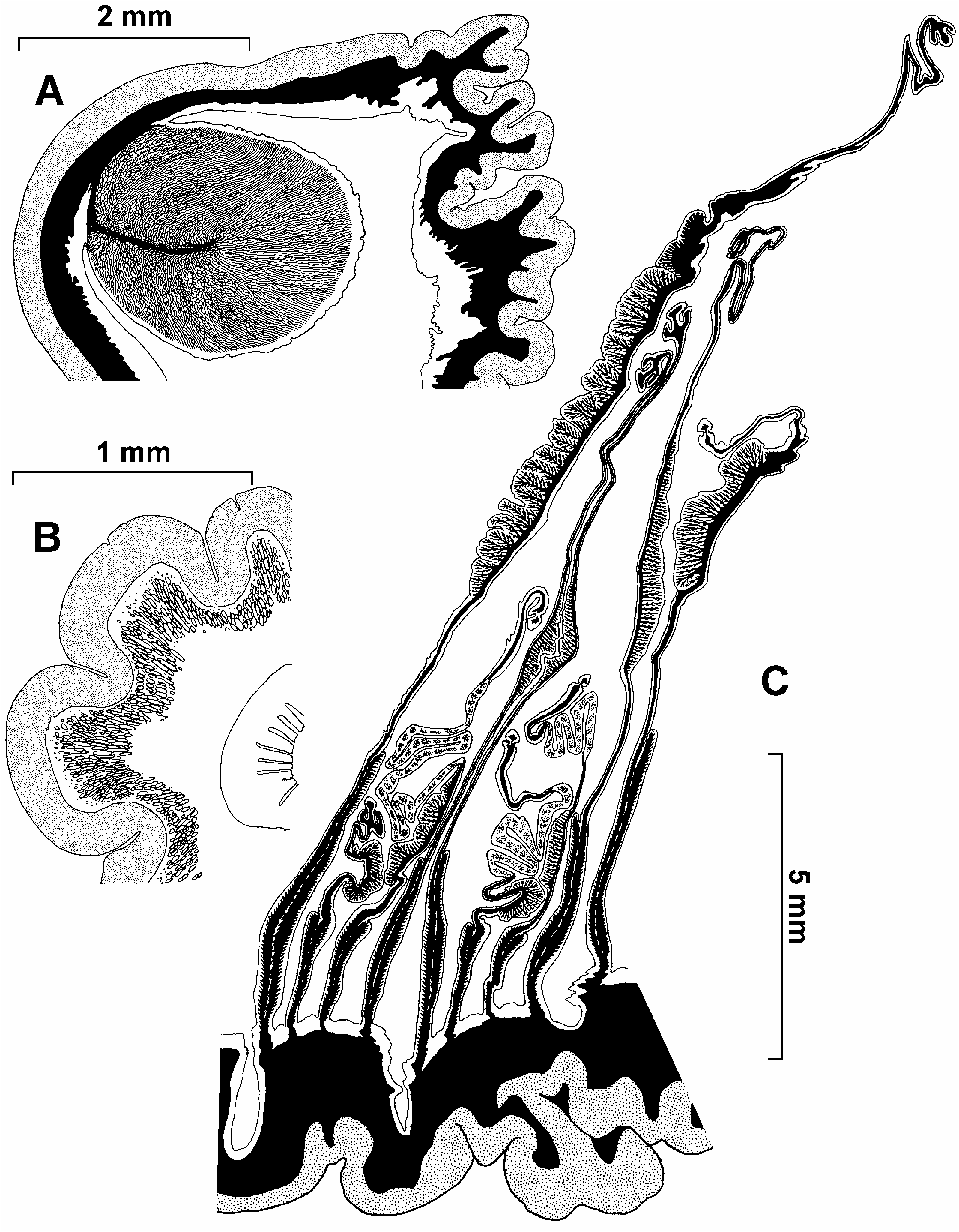
Internal structure. The mesogloea of the column is thick, up to 4 mm in large specimens. The columnar verrucae on the transverse sections appear as places with thin mesogloea between the deep tubular evagination of the endoderm and invagination of the ectoderm. Although the presence of the verrucae may not be evident on the contracted specimens, their presence can be always detected in sections.
The marginal endodermal sphincter is strong, circumscribed, with one short main lamella ( Figure 9A View Figure 9 ). Radial muscles of the oral disk and longitudinal muscles of the tentacles are mesogloeal ( Figure 9B View Figure 9 ). Two deep siphonoglyphs are supported by directives.
The first 12 pairs of mesenteries are equally developed as belonging to the same cycle. Accordingly, distribution of the mesenterial pairs may be described as 12+12+24+49 (specimen 238/10). Some specimens may be perfectly 11-merous (11+11+22+45, specimen 231/8) or 13-merous (13+12+26+50, specimen 260/14), 14-merous (14+14+28+52, specimen 67/12) or, often, the arrangement of the mesenteries may be irregular with a different number of mesenterial pairs on the left and right sides of the directive plane. The number of mesenteries appears to be somewhat greater than the number of tentacles (e.g. 186 tentacles and 202 mesenteries in specimen 260/14, or 174 tentacles and 216 mesenteries in specimen 67/12). Gonads are present only on the two last cycles of the mesenteries, the first two cycles (24 pairs) are always sterile. Oocytes to 0.8 mm and spermatic vesicles to 0.15 mm in diameter. The retractor muscles are rather weak and diffuse to restricted. Welldeveloped parietobasilar muscles form a free flap. Parietal muscles on transverse sections extend from the column wall to halfway to retractors ( Figure 9C View Figure 9 ).
The sexes are separate, no embryos were found in any specimen.
Size and distribution of cnidae (letters in brackets refer to Figure 10 View Figure 10 , all measurements in Mm; N is the proportion of examined specimens that had a particular type of cnida; distribution of all cnidae is confirmed on sections):
Column basitrichs (a): 6–12×1.5–2.5 (common), N54/7
basitrichs (b): 14–29×2–3.5 (numerous), N57/7
p-mastigophores A (c): 16–24×5–6 (very rare), N54/7
Tentacles spirocysts (a): 21–55×2–4 (numerous), N56/6 basitrichs (b): 8–14×1.5–2.5 (common, mostly crescent-shaped), N56/6 basitrichs (c): 18–32×2–3.5 (numerous), N56/6 p-mastigophores A (d): 19–22×5 (very rare), N55/6
Actinopharynx basitrichs (a): 52–97×4.5–7 (numerous), N58/8 p-mastigophores A (b): 27–38×5–6 (very rare), N55/8
Filaments basitrichs (a): 12–21×2–2.5 (common), N59/10 basitrichs (b): 51–81×5–6.5 (common), N510/10 p-mastigophores A (c): 21–38×5–6 (common), N510/10 p-mastigophores B (d): 29–51×5–8 (numerous), N510/10
Presence of rare p-mastigophores A in the ectoderm of the tentacles appears to be a characteristic feature of the species (these cells were not detected in the tentacles of the other examined species of Urticina and Cribrinopsis ).
Habitat
The specimens were found from 3 to 25 m depth, and never in the intertidal zone in the studied area. They are always attached to large boulders or rock, and are never buried in sand. The column is always clear from gravel particles or other foreign matter. Several large specimens were recorded attached around the den of a large octopus.
Etymology
The species is named after Dr Sergey Grebelnyi who first recognised this species in 1983.
Remarks
The species is characterised by its large size and numerous non-adhesive ampullaceous verrucae spread over the whole column. They form a distinct annulus on the margin and may become large and inflated when the specimen contracts. The species is most closely related to the northern U. eques . Verrucae in the latter species are weakly developed, and usually the mesenteries are arranged decamerously ( Stephenson 1935), while not one of the numerous dissected specimens of the present material has decamerously arranged mesenteries. Also, there are some differences in the cnidom, which seem to be real rather than apparent. In particular, U. grebelnyi sp. nov., unlike U. eques , has p-mastigophores A in the ectoderm of the column and tentacles and crescent-shaped basitrichs in the tentacles, and appears to have only one type of small basitrich in the filaments, while U. eques has two types of small basitrichs in the filaments. This distinction, however, needs to be confirmed on material from different locations.
The present species is not related to U. felina and U. crassicornis , the former having strongly adhesive verrucae (very different from the non-adhesive verrucae of U. grebelnyi sp. nov.) and the latter always has a smooth column. However, it is certainly conspecific with the specimens from Puget Sound identified as Tealia crassicornis by Chia and Spaulding (1972) and Sebens and Laakso (1977) (see Remarks, U. crassicornis ).
Urticina tuberculata: Zamponi and Acuna, 1996 View in CoL from Vancouver Island appears to be conspecific with the present species. Most authors who studied U. tuberculata View in CoL in Europe considered it to be very similar to U. felina View in CoL and ‘‘perhaps hardly worthy of varietal rank’’ ( Stephenson 1935, p 143), and Manuel (1981) finally synonymised them. Indeed, according to the original description of Actinia tuberculata View in CoL cited by Gosse (1860, p 217), the body is ‘‘densely covered with large grayish-white tubercles, the apex of each tubercle is depressed’’. The depressions in these tubercles suggest that they are true adhesive verrucae similar to those of U. felina View in CoL . Non-adhesive verrucae of U. grebelnyi View in CoL sp. nov. may appear as depressed, but only on the preserved specimens, while the original description of A. tuberculata View in CoL was made from a living specimen.
Urticina piscivora ( Sebens and Laakso 1977) View in CoL differs in possessing verrucae only in the upper portion of the column where they are arranged in about five rows and, in addition, the colour is different. Urticina columbiana Verrill, 1922 View in CoL and U. kurila Averincev, 1967 View in CoL , unlike the present species, have well-developed adhesive verrucae and live buried in sand.
This spectacular species is very abundant in some localities in the vicinity of the Petropavlovsk-Kamchatsky and is the largest actiniid species recorded on the depths accessible for divers in the studied area. It certainly is present in Puget Sound.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Urticina
| Sanamyan, N. P. & Sanamyan, K. E. 2006 |
Urticina tuberculata : Zamponi and Acuna 1996 , p 3
| Zamponi MO & Acuna FH 1996: 3 |
Urticina eques :
| Manuel RL 1981: 109 |
Urticina felina :
| Manuel RL 1981: 106 |
Tealia crassicornis : Chia and Spaulding 1972 , p 206
| Sebens KP & Laakso G 1977: 165 |
| Widersten B 1976: 865 |
| Chia FS & Spaulding J 1972: 206 |
Tealia crassicornis : Chia and Spaulding 1972 , p 206
| Sebens KP & Laakso G 1977: 165 |
| Chia FS & Spaulding J 1972: 206 |
Tealia crassicornis :
| Hand C 1955: 72 |
Tealia lofotensis : Hand 1955 , p 80
| Sebens KP & Laakso G 1977: 162 |
| Hand C 1955: 80 |
Tealia coriacea : Hand 1955 , p 76
| Hand C 1955: 76 |
Tealia felina var. crassicornis :
| Stephenson TA 1935: 150 |
Tealia felina var. lofotensis : Stephenson 1935 , p 147
| Stephenson TA 1935: 147 |
Tealia felina var. coriacea :
| Stephenson TA 1935: 140 |
Urticina felina crassicornis :
| Carlgren O 1921: 170 |
Urticina felina lofotensis :
| Carlgren O 1921: 168 |
Urticina felina coriacea
| Carlgren O 1921: 163 |
Urticina felina :
| McMurrich JP 1911: 65 |
Tealia lofotensis :
| Carlgren O 1902: 42 |
Madoniactis lofotensis
| Danielssen DC 1890: 47 |
Bolocera eques
| Gosse PH 1860: 351 |
Actinia tuberculata
| Cocks WP 1850: 94 |
Actinia tuberculata
| Cocks WP 1850: 94 |
Actinia coriacea
| Cuvier G 1798: 653 |
Actinia crassicornis Müller 1776 , p 231
| Muller OF 1776: 231 |
Actinia crassicornis Müller 1776 , p 231
| Muller OF 1776: 231 |
Priapus felinus
| Linnaeus AC 1761: 510 |