
Lophochaeta ignota Štolc, 1886
|
publication ID |
https://doi.org/10.5281/zenodo.193114 |
|
DOI |
https://doi.org/10.5281/zenodo.6211157 |
|
persistent identifier |
https://treatment.plazi.org/id/C8764570-FFAD-AA0F-71C8-BE08FC82F868 |
|
treatment provided by |
Plazi |
|
scientific name |
Lophochaeta ignota Štolc, 1886 |
| status |
|
Lophochaeta ignota Štolc, 1886 View in CoL
( Figures 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 )
Lophochaeta ignota Štolc, 1886
Lophochaeta ignota Štolc : Štolc 1888
Tubifex filum Michaelsen, 1901
Tubifex longiseta Bretscher, 1905
Tubifex ignotus (Štolc) : Laakso 1969. Hrabë 1981. Chekanovskaya 1981 For references before 1971 see Brinkhurst and Jamieson 1971
Examined material. From Apraiz karstic spring ( 6 September 2005), UTM coordinates X:524225, Y:4801350, Z:22. One whole-mounted in lactophenol, 3 sectioned, 4 dissected (one of them not fully mature) and 24 specimens preserved in alcohol 70% (at different stages of maturity). From Iturgoien cave ( 6 September 2005) one dissected specimen not fully mature.
Localities. Apraiz spring and Iturgoien cave (X:524217, Y:4801296, Z:21) both are very close and connected subterranean sites in Busturia (Santa Eufemia–Ereñozar karst unit, Biscay, Spain) ( 6 September 2005).
Description. Long worms, body length of preserved mature specimens up to 40 mm with 160 segments; width in segment VIII 0.3–0.4 mm; and down to 0.15 mm in the posterior region of the body.
Prostomium rounded, 120–135 µm long and 115–145 µm wide. Body wall consisting of a muscular layer 5–7 µm thick, and an epidermal layer 5–10 µm thick covered by a cuticle of 2–3 µm. Anterior region of the body with secondary annulation; in two examined mature individuals it was apparent from segment II to V and II to VIII, respectively. Clitellum in segments XI–XII, not much elevated. One pair of spermathecal pores in X, close and lateral to the ventral bundles of chaetae. When completely mature (with eggs into the egg sac), one pair of male pores open close to ventral bundles of chaetae, in a hollow formed in the ventral side of segment XI.
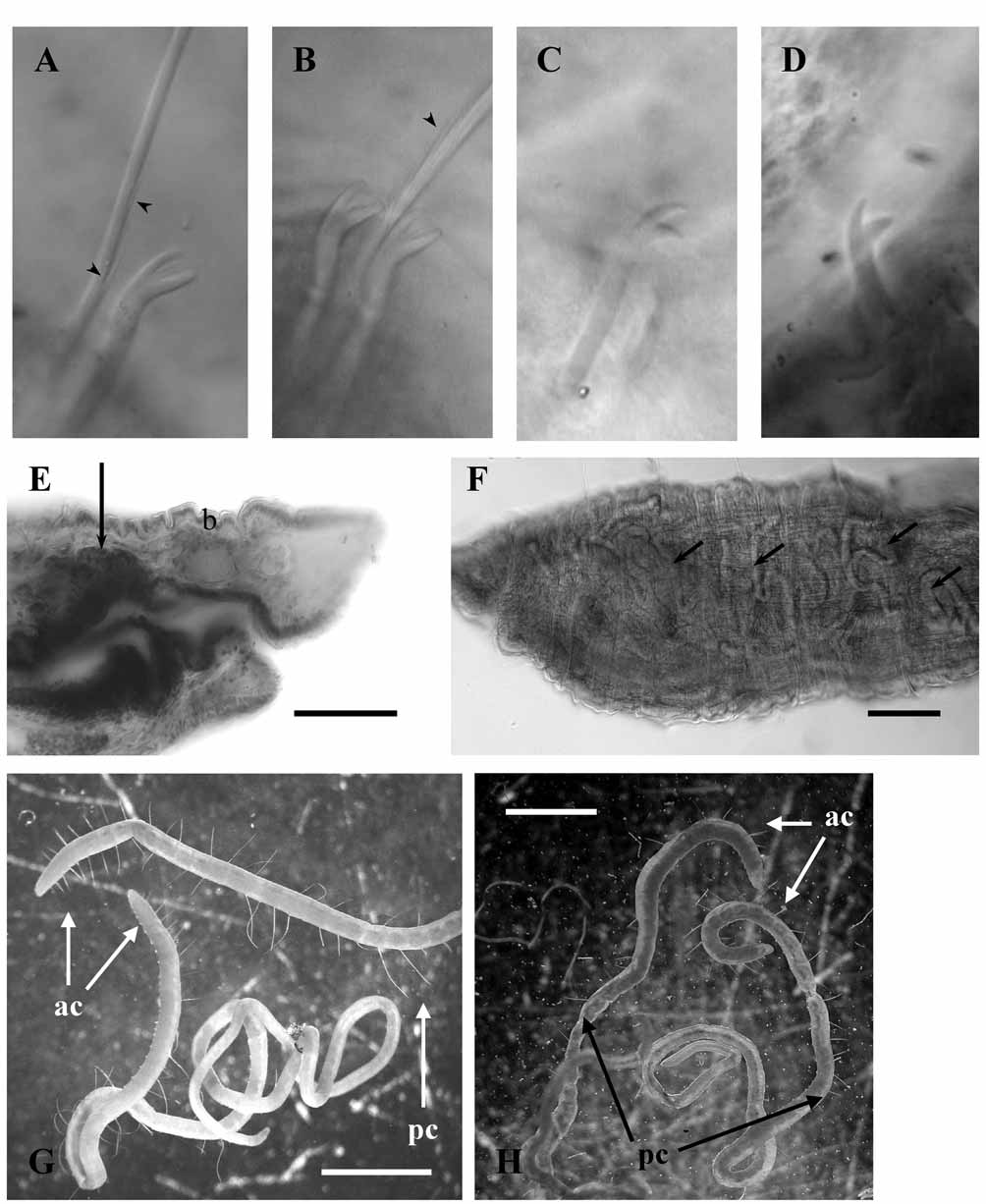
Dorsal bundles of chaetae consist of hair and pectinate chaetae. Hair chaetae very long and hispid ( Fig. 7 View FIGURE 7 A, B) covered by fine bristles visible under 400x magnification. In preclitellar segments 1–2 hairs per bundle, one in postclitellar segments and absent at the hind part of the body. Length of hair chaetae grows backwards, up to 350 µm long in segment III; up to 480 µm in V; up to 650 µm in VIII (approx. 2 times the body diameter) and up to 850 µm (2 to 3 times the body diameter) in postclitellar segments. In preclitellar dorsal bundles, 2–3 characteristic pectinate chaetae per bundle, 88–95 µm long, with distal nodulus, the proximal part of the chaeta straight and the distal part curved; stout teeth up to 7 µm long, with 2 intermediate thin teeth of about equal length ( Fig. 7 View FIGURE 7 A, B)). In posterior segments, one pectinate chaeta per bundle, 60–78 µm long, with shorter teeth and gradually losing the characteristic curved shape of the distal part of the chaeta towards the hind region of the body, where regular sigmoid chaetae without intermediary teeth are found. Ventral bundles in anterior segments with 3–4 bifid chaetae (52–82 µm long), distal tooth longer than proximal one ( Fig. 7 View FIGURE 7 C); in postclitellar segments 2–3 bifid chaetae, 68–73 µm long, with distal tooth shorter and thinner than proximal one ( Fig. 7 View FIGURE 7 D). No modified genital chaetae.
Brain back to intersegment 1/2. Coelomocytes not visible within the coelomic cavity in most specimens, although in some occasions small ones ( ca 3 µm diameter) were observed. Typical muscular dorsal pharyngeal pad not developed. Pharyngeal glands from segment III back to segment VI, dorsal to the pharynx in segment III ( Fig. 7 View FIGURE 7 E), lateral and ventral to the gut in IV and V, and ventral in VI. A layer of chloragogen cells on the digestive tract, from segment VI, backwards. Oesophagus opens gradually into the intestine. Dorsal and ventral vessels very apparent throughout the body and commisural vessels noticeable in anterior segments ( Fig. 7 View FIGURE 7 F).
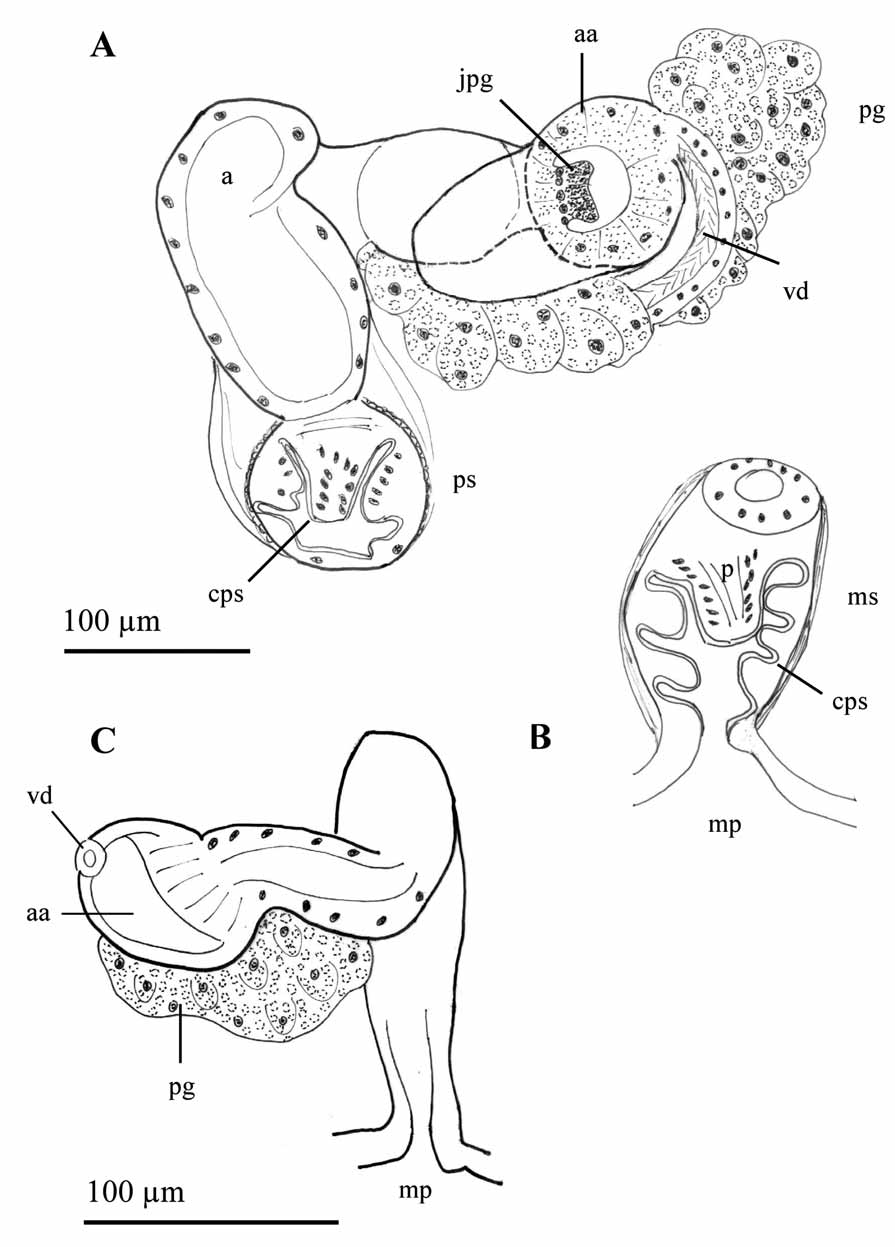
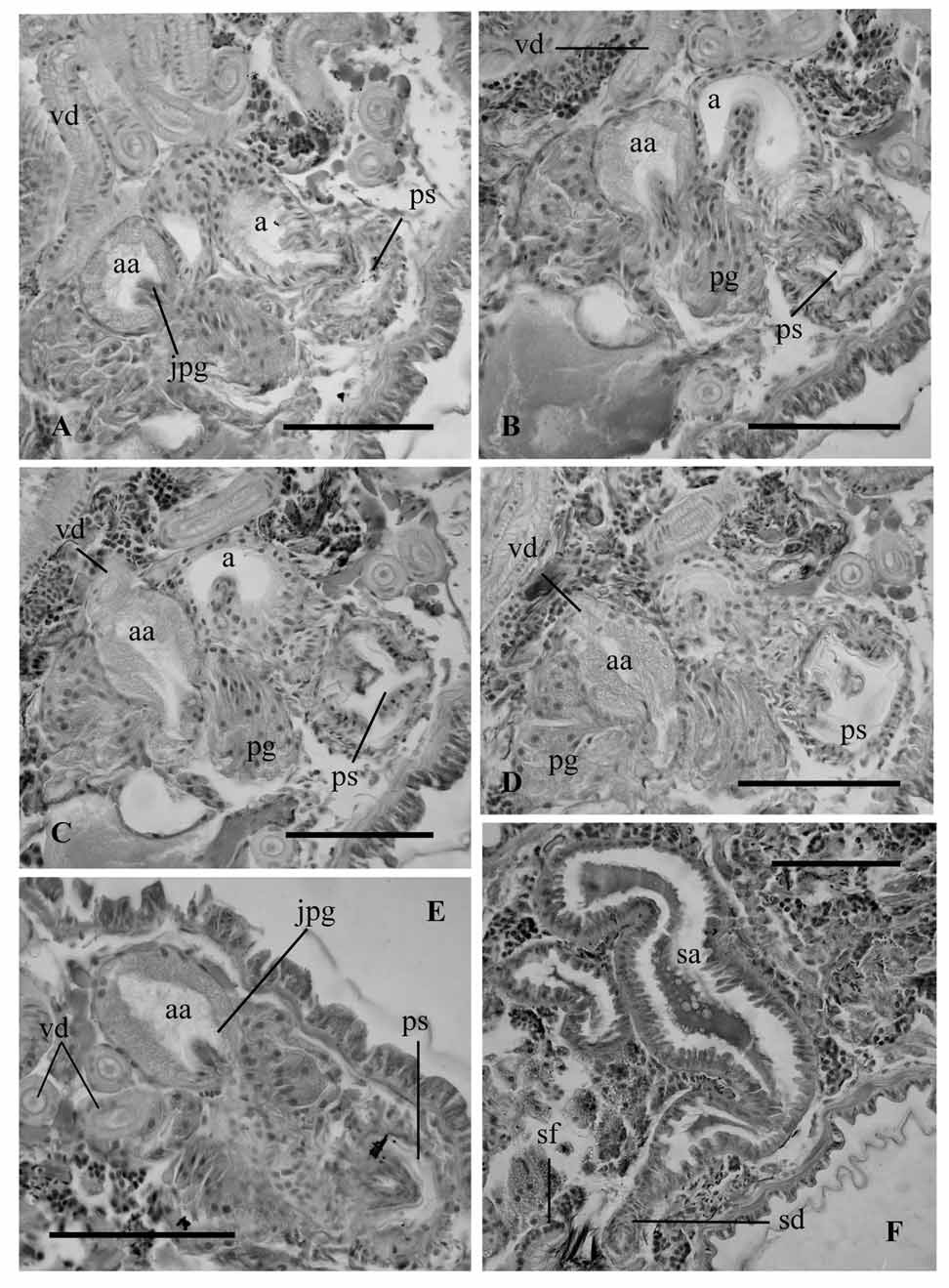
One pair of testes in X and one pair of ovaries in XI. Sperm sacs developed anteriorly to segment IX and back to segment XIII. Egg sac can reach segment XV. All following measurements on the male duct were obtained from egg-bearing individuals. Male funnel 100–107 µm high, attached to the septum in 10/11. Very long vas deferens, forming numerous loops in segments XI and XII, ciliated and about the same diameter throughout its length, varying in different specimens from 30 to 40 µm (in one specimen briefly narrowing just before the junction with the sperm funnel); epithelial layer 7.5–10 µm thick. Atria tubular in segment XI, approximately 275–325 µm long, 34–43 µm diameter, and covered by a very thin muscular layer. Maximum diameter of atrium up to 60 µm in the distal portion; atrium folds several times, and enters the penial sac through a narrow section (ca. 30 µm diameter). Atrium expands entally, forming an oval ampulla (70–80 µm wide, and 105–150 µm long) ( Fig. 8 View FIGURE 8 A). Atrial epithelium simple in the ectal section, and glandular (finely granular) in the ental ampulla. Epithelial cells at the atrial ampulla higher (up to 25 µm) than those in the other sections of the atrium (13–18 µm). Vas deferens enters the distal part of the atrium opposite to the junction of the compact prostate gland ( Fig. 9 View FIGURE 9 A–D). Atrium opens into elongated penial sac, 100 µm wide, reinforced by both longitudinal and circular musculature. In fully mature specimens, the internal walls of the penial sacs are covered by a thin cuticle, and have several lateral folds ( Fig. 8 View FIGURE 8 B and 9B–E). Short, conical penis (up to 37 µm long and 37 µm maximum diameter) is lined with a cuticular layer thinner than that of the body wall; its outline follows the shape of the penis. The penis is not free within this cuticular sheath. Female funnels were not observed.
One pair of spermathecae in segment X. Spermathecal ampulla elongated, up to 425 µm long and 132 µm diameter in the examined specimens, gradually narrowing towards the spermathecal duct, which is relatively short (50–70 µm long and 25 µm wide) ( Fig. 9 View FIGURE 9 F). Spermathecal epithelium up to 10 µm high. Undetermined matter fills the lumen of the ampulla, with a smooth uniform surface that resembles a liquid, and no remnants of spermatozeugmata or spermatozoids were observed.
Distribution and habitat. The species has been widely reported in lakes and rivers of the Holarctic region, Africa ( Tunisia and South Africa) and South America (Lake Titicaca). In the Iberian Peninsula, it has been reported in epigean waters, usually immature; in karstic springs of the study area it has been found mature for the first time.
Taxonomic remarks. The description of the new material of the species agrees with the diagnosis of Lophochaeta ignota by Štolc (1886, 1888), whose description was meticulous on nerve and vascular systems but poor on the reproductive organs. Štolc did not measure the length of the hair chaetae but mentioned their hispid nature. The new material also agrees with the diagnosis of Lophochaeta by Holmquist (1985), being remarkable the presence of hispid (barbed) hair chaetae; the tubular shape of the atrium, with an ental ampulla, where prostate and vas deferens enter opposite each other; and the long and throughout ciliated vas deferens. The position of spermathecal pores, lateral in Tubifex and ventral in Lophochaeta , could be a character subject to variation at the species level, although Holmquist introduced it in the diagnosis of the genera Tubifex (as lateral) and Lophochaeta (as ventral). This variability can also be applied for the shape of spermathecal ampulla (rounded vs. sac-like) or for the length of the spemathecal duct (short vs. long).
The synonymy of Tubifex filum with Lophochaeta ignota (proposed by Hrabë 1962, and widely accepted) is admitted by the authors, although we recognize that there are some blank gaps in the original description of the species. Thus, Štolc (1886, 1888) did not mention the presence of exceptionally long hair chaetae in the diagnosis of Lophochaeta ignota although this character was stated in subsequent descriptions of the species ( Michaelsen 1901, Hrabë 1962). By other side, Tubifex filum was described by Michaelsen (1901) as a long, thin and threadlike worm, with 2 (3) very fine and long hair chaetae with maximum length behind the clitellum (0.9 mm), but there was no mention to barbs. According to our measurements in North European populations of L. ignota , the length of the hair chaetae can be variable (up to 550 µm in postclitellar segments, in the Russian population; up to 1350 µm in postclitellar segments, in the Estonian population) ( Fig. 7 View FIGURE 7 G, H). Regarding to the barbs on the hair chaetae, all examined specimens from different geographic areas have hispid chaetae, which were observed at a magnification of 400x and with interferential contrast.
The general description of the male duct of the species L. ignota by Hrabë (1962, 1981, as Tubifex ignotus ) mostly agrees with ours, except that he states that the vas deferens gradually dilates from the ental (22 µm diameter) to the ectal part (32–40 µm diameter). He also described the junction of vas deferens with the atrium as apical, in the small spherical ampulla of the atrium. It is interesting to note that this general sketch of the atrium is similar to that found in individuals which have not formed eggs yet (see Fig. 8 View FIGURE 8 C). Our examined specimens have glandular epithelium mostly restricted to the apical ampulla while atrial walls were described as “glandular for the most part” by Chekanovskaya (1981: 340). She also stated that atrium “posteriorly narrows abruptly to 44 µm”, probably referring to the narrowing that occurs just before entering the penial sac.
Hrabë (1981) described the penis of L. ignota (as T. ignotus ) being cylindrical and covered by a strong cuticle, but not forming a cuticular tube of permanent shape. In our specimens, the cuticle although well visible, does not appear strong and it conforms to the shape of the penis, which is not free inside a cuticular sheath. This can be interpreted in the same way as Hrabë’s description. Chekanovskaya (1981) described the penis of the species as “small and rounded” within a folded penial sac. In the examined specimens from our collection, the penis is conical and short in relation to the length of the penial sac, which is also folded ( Fig. 8 View FIGURE 8 B).
The general shape and structure of the spermatheca in present description agrees with the description given by Štolc (1886, 1888), Hrabë (1962, 1981) and Chekanovskaya (1981). Spermatozeugmata were only mentioned by Hrabë (1962) but drawings or detailed descriptions were lacking. The examination of specimens from Russia revealed the presence of sperm into the spermathecal ampulla, but without forming spermatozeugmata (observation confirmed by M. Ferraguti).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lophochaeta ignota Štolc, 1886
| Rodriguez, Pilar & Achurra, Ainara 2010 |
Tubifex longiseta
| Bretscher 1905 |
Tubifex filum
| Michaelsen 1901 |
Lophochaeta ignota Štolc, 1886
| Stolc 1886 |