
Chorthippus apricarius (Linnaeus, 1758)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4318.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:A9Ee2696-C9A3-4C19-984D-Bbf44129C7C9 |
|
DOI |
https://doi.org/10.5281/zenodo.6002062 |
|
persistent identifier |
https://treatment.plazi.org/id/BF2187E3-FFA2-484E-FF06-4FC0FACDE167 |
|
treatment provided by |
Plazi |
|
scientific name |
Chorthippus apricarius (Linnaeus, 1758) |
| status |
|
8. Chorthippus apricarius (Linnaeus, 1758)
Figs. 61–85 View FIGURES 61 – 74 View FIGURES 75 – 85
Ch. apricarius asiaticus Mistshenko, 1951 , syn. n.
Distribution. Europe, including European Russia, the Caucasus, Kazakhstan, Central Asia, South Siberia up to Trans-Baikal Kray and Southeast Sakha (Yakutia), Mongolia, and North China. Separate subspecies are described from Ciscaucasia and the Caucasus ( Ch. apricarius major Pylnov, 1914 , Ch. apricarius caucasicus Mistshenko, 1951 , and Ch. apricarius ciscaucasicus Mistshenko, 1951 ), and from the mountains of Central Asia ( Ch. apricarius asiaticus Mistshenko, 1951 ).
Localities. Ch. apricarius apricarius :
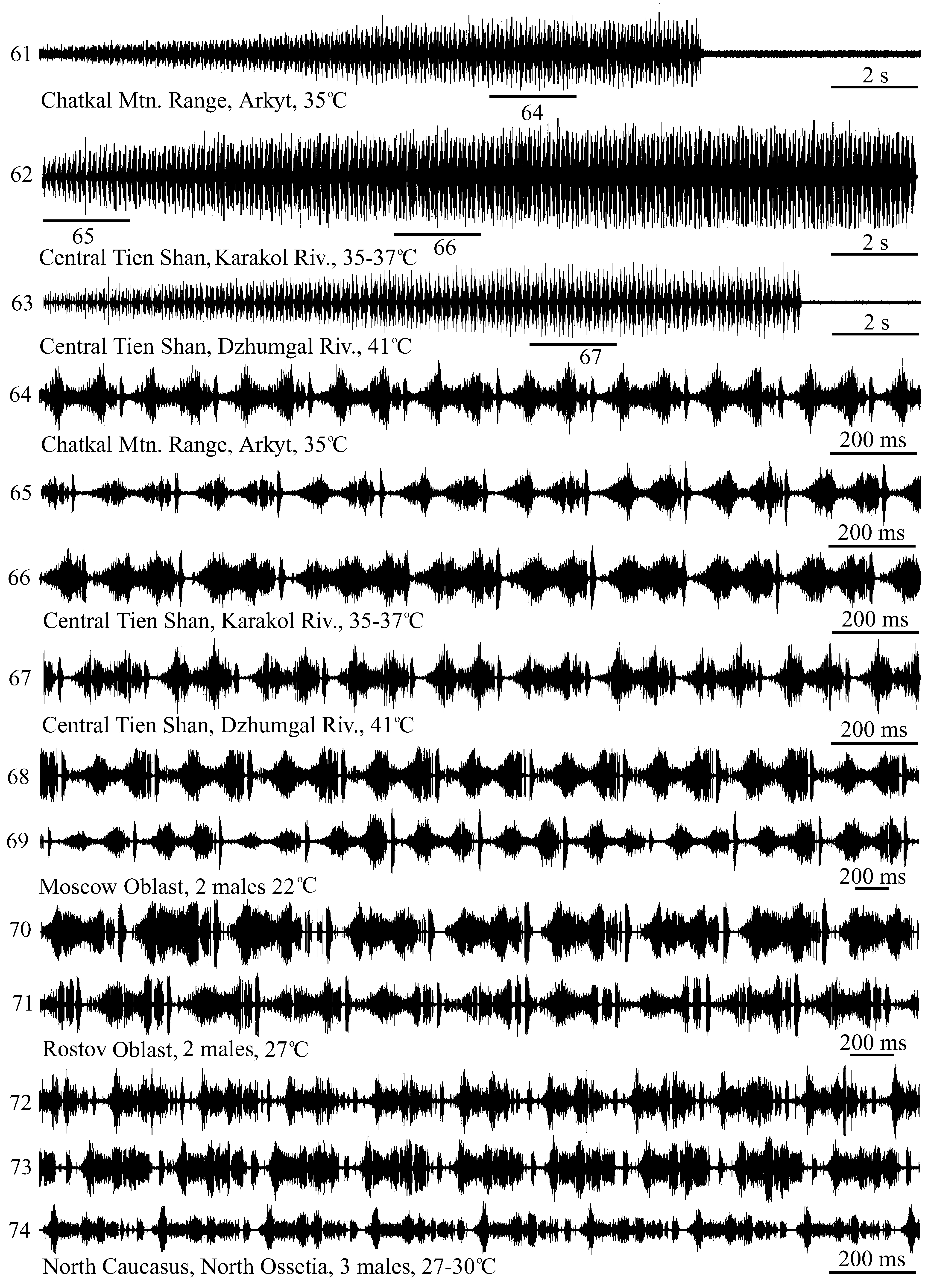
3. The West Tien Shan Mts. , Chatkal Mtn. Range, Sary Chelek Nature Reserve, environs of Arkyt Village, glades in a fruit-walnut forest, 21. VII. 2008. Signals of 5 ♂ recorded on disk at 30 and 35–40o C ( Figs. 61 and 64 View FIGURES 61 – 74 ).
4. The Central Tien Shan Mts. , the Kekemeren River Basin , the Western Karakol River ca. 10 km from the mouth (10 km East-North-East from Suusamyr Village), mountain steppes on the right bank, 6. VII. 2016. Signals of 1 ♂ recorded on disk at 35–37o C ( Figs. 62 and 65–66 View FIGURES 61 – 74 ).
6. The Central Tien Shan Mts., the Dzhumgal River Valley East of Chaek , 30.VI.2014. Signals of 1 ♂ recorded on disk at 41o C ( Figs. 63 and 67 View FIGURES 61 – 74 ).
Moscow Oblast, Voskresensk District, environs of Beloozerskiy Town , 8. VII. 1992 (not shown on the map). Signals of 3 ♂ recorded on tape at 22o C ( Figs. 68–69 View FIGURES 61 – 74 ).
Rostov Oblast, Oblivskiy District, environs of Sosnovy (= Oporny ) Village on the Chir River , 16. VIII. 1992 (not shown on the map). Signals of 2 ♂ recorded on tape at 27o C ( Figs. 70–71 View FIGURES 61 – 74 ).
The North Caucasus, North Ossetia, the Ardon River Basin , Tsey Mtn. Range, environs of the Upper Tsey Village , 1700–1800 m above sea level, 27. VII. 1990 (not shown on the map). Signals of 3 ♂ recorded on tape at 27–30o C ( Figs. 72–74 View FIGURES 61 – 74 ).
References to song. Ch. apricarius apricarius (Linnaeus, 1758) . Ragge & Reynolds (1998): recordings from Western Europe; Vedenina & Bukhvalova (2001): recordings from the Ukraine, central and southern parts of European Russia, South Kazakhstan, and Altai Mts.; Benediktov (2005): recordings from Altai Mts.; Iorgu & Iorgu (2011): recordings from Romania. Ch. apricarius major Pylnov, 1914 . Vedenina & Bukhvalova (2001): recordings from the North Caucasus, North Ossetia.
Song. Male calling song is a prolonged echeme-sequence lasting about 15–30 s. The song begins quietly and gradually reaches maximum intensity in the first third or half of its duration ( Figs. 61–63 View FIGURES 61 – 74 ). Typically, each echeme consists of an abrupt “click” followed by two syllables ( Figs. 64–71 View FIGURES 61 – 74 ). Echeme repetition period gradually decreases towards the middle of a song and more abruptly increases in last ca. 10 echemes ( Tishechkin & Bukhvalova, 2009b, fig. 84). In males from Kyrgyzstan echeme repetition period in the middle of a song averages 180–240 ms ( Figs. 64–67 View FIGURES 61 – 74 ). In the nominotypical subspecies, there are few if any gaps in syllables ( Figs. 64–71 View FIGURES 61 – 74 ), whereas in Ch. apricarius major typically, several gaps present in each syllable ( Figs. 72–74 View FIGURES 61 – 74 ).
In contrast to some other species, rhythmically repeated elements in the songs of Ch. apricarius and G. mollis are produced by several up- and downstrokes of hind legs, i. e. formally should be referred to as echemes. However, on oscillograms they look like syllables in the songs of other species (e. g. in S. fischeri , G. sibiricus , G. maritimus ). For this reason, for the purposes of comparative signal analysis it is sometimes more convenient to refer such elements to as syllables despite the differences in leg movements ( Tishechkin & Bukhvalova, 2009b).
Comparative notes. The Caucasian subspecies, Ch. apricarius major distinctly differs from Ch. apricarius apricarius in more complicated echeme pattern ( Figs. 72–74 View FIGURES 61 – 74 ), although is similar with it in general pattern of the song and in echeme repetition period ( Vedenina & Bukhvalova, 2001). On the other hand, we have not found any differences in signal patterns between Ch. apricarius from Kyrgyzstan ( Figs. 64–67 View FIGURES 61 – 74 ; Ch. apricarius asiaticus , according to Bey-Bienko & Mistshenko, 1951) and from European Russia ( Figs. 68–71 View FIGURES 61 – 74 ; the nominotypical subspecies, according to Bey-Bienko & Mistshenko, 1951).
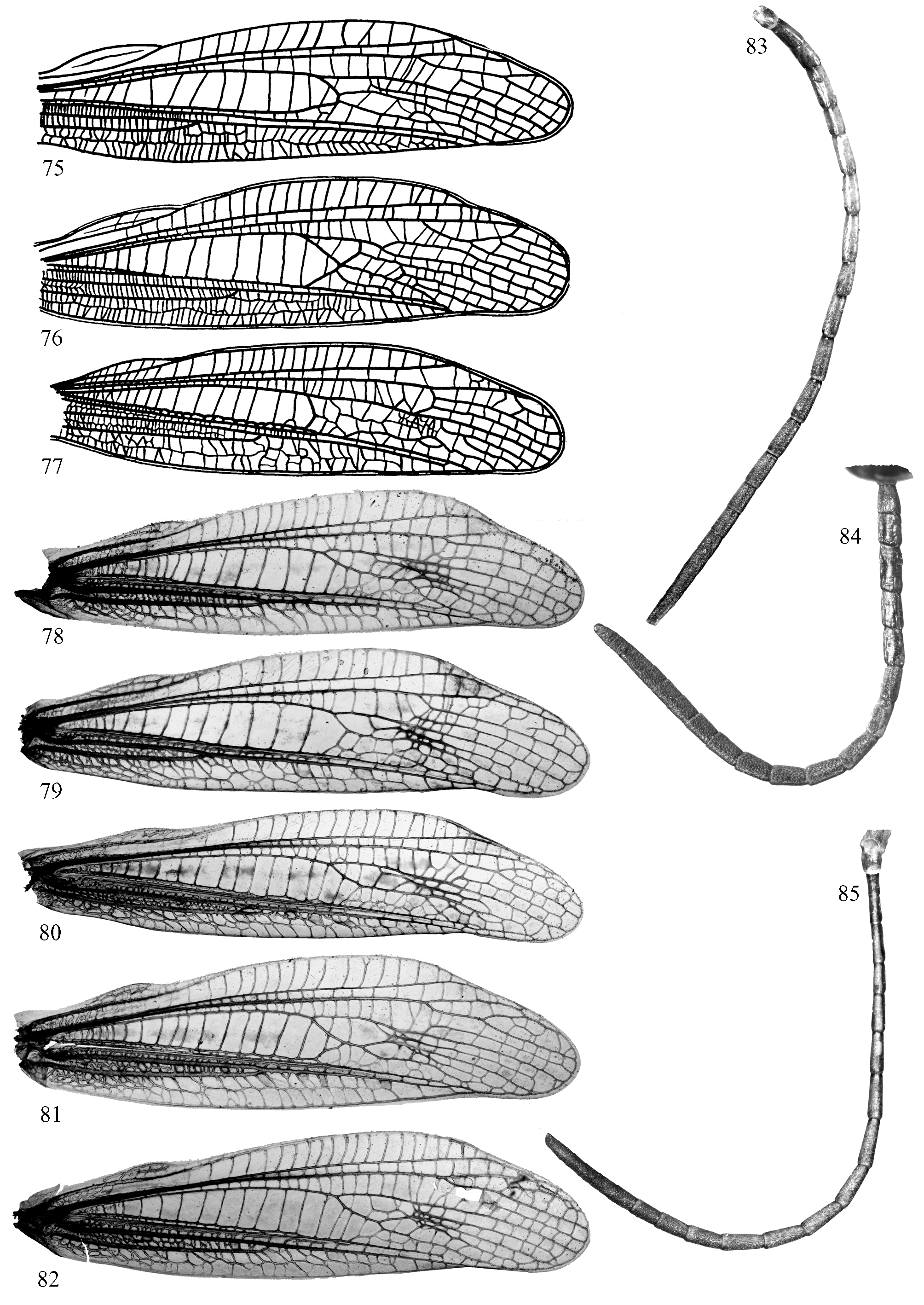
According to original description (Bey-Bienko & Mistshenko, 1951), in Ch. apricarius asiaticus cubital veins on forewing are separated throughout all the length of medial area so that cubital area is always distinct ( Fig. 75 View FIGURES 75 – 85 , holotype, after Bey-Bienko & Mistshenko, 1951) and antennae are rather thick (length to width ratio of a middle antennal segment averages 1.25–1.75). In the nominotypical subspecies cubital veins on forewing are separated only at the base ( Fig. 76 View FIGURES 75 – 85 ); occasionally, they are separated throughout all the length of medial area ( Fig. 77 View FIGURES 75 – 85 ), but in this case antennae are thin (length to width ratio of a middle antennal segment averages 2–2.25).
In males from Kyrgyzstan, which signals were studied, cubital veins on forewing are separated only at the base (males from the Central Tien Shan Mts., Figs. 78–79 View FIGURES 75 – 85 ), or form distinct but very narrow cubital area (males from the West Tien Shan Mts., Figs. 80–82 View FIGURES 75 – 85 ). Proportions of middle antennal segments vary greatly even in males from the same locality collected at the same date. Photographs of forewings and antennae of three males from Sary Chelek Nature Reserve collected one after another on the same glade are given on Figs. 80–85 View FIGURES 75 – 85 ; signals of all males were recorded on disk. As can be seen from these photographs, two males should be attributed to the nominotypical subspecies ( Figs. 80, 83 and 82, 85 View FIGURES 75 – 85 : combination of distinct cubital area and thin antennae), and one to Ch. apricarius asiaticus ( Figs. 81 and 84 View FIGURES 75 – 85 : distinct cubital area but thick antennae). Thus, all diagnostic traits mentioned in original description actually cannot be used for discrimination between Ch. apricarius apricarius and Ch. apricarius asiaticus , and both forms can be found in the mountains of Kyrgyzstan. For this reason, we establish a synonymy Ch. apricarius apricarius (Linnaeus, 1758) = Ch. apricarius asiaticus Mistshenko, 1951 , syn. n.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Gomphocerinae |
|
Genus |
Chorthippus apricarius (Linnaeus, 1758)
| Tishechkin, Dmitri Yu. 2017 |
Ch. apricarius asiaticus
| Mistshenko 1951 |
Chorthippus apricarius apricarius sensu
| Bey-Bienko & Mistshenko 1951 |
Ch. apricarius major
| Pylnov 1914 |
Ch. apricarius major
| Pylnov 1914 |
Ch. apricarius major
| Pylnov 1914 |
Ch. apricarius
| apricarius (Linnaeus 1758 |
Ch. apricarius apricarius
| apricarius (Linnaeus 1758 |