
Mycetophyllia, MILNE EDWARDS & HAIME, 1848: 491 - 492
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2012.00855.x |
|
persistent identifier |
https://treatment.plazi.org/id/B26DA91C-6D13-172E-E045-FBB8D2AAFE6D |
|
treatment provided by |
Marcus |
|
scientific name |
Mycetophyllia |
| status |
|
GENUS MYCETOPHYLLIA MILNE EDWARDS & HAIME, 1848: 491–492 View in CoL ( FIGS 9E, F View Figure 9 , 14A–J View Figure 14 , 19G–O View Figure 19 , 23G–O View Figure 23 )
Synonyms: None.
Type species: Mycetophyllia lamarckiana Milne Edwards & Haime, 1849: 258, holotype = MNHN- Scle910 ( Fig. 9E, F View Figure 9 ); by subsequent designation, Milne Edwards & Haime, 1849: 258.
Type species locality: ‘Patrie inconnue’ ( Milne Edwards & Haime, 1849: 258) [Recent].
Original description: ’Très-voisin des Symphyllies, mais n’offrant pas de columelle et ayant les calices très-peu profonds. Murailles peu marqués, et cloisons en petit nombre.’ ( Milne Edwards & Haime, 1848: 491).
Subsequent morphological descriptions ( Matthai, 1928 and later): Matthai, 1928: 249–255; Vaughan & Wells (1943: 193, 196); Wells (1956: F419); Wells (1973: 34–43); Walton Smith (1971: 93–94); Zlatarski & Estalella (1982: 182–224); Veron (2000: vol. 3: 72–79).
Diagnosis: Macromorphology: colonial. Intracalicular and/or circumoral budding. Meandroid (uniserial and multiserial), with and without collines; centres spaced 5–15 mm apart; < three septal cycles, equal; limited ·
coenosteum and incomplete corallite walls, mostly confluent septa; absent or feeble discontinuous columella with lamellar linkage; reduced epitheca; paliform lobes; abundant endotheca ( Figs 9E, F View Figure 9 , 14A–J View Figure 14 ).
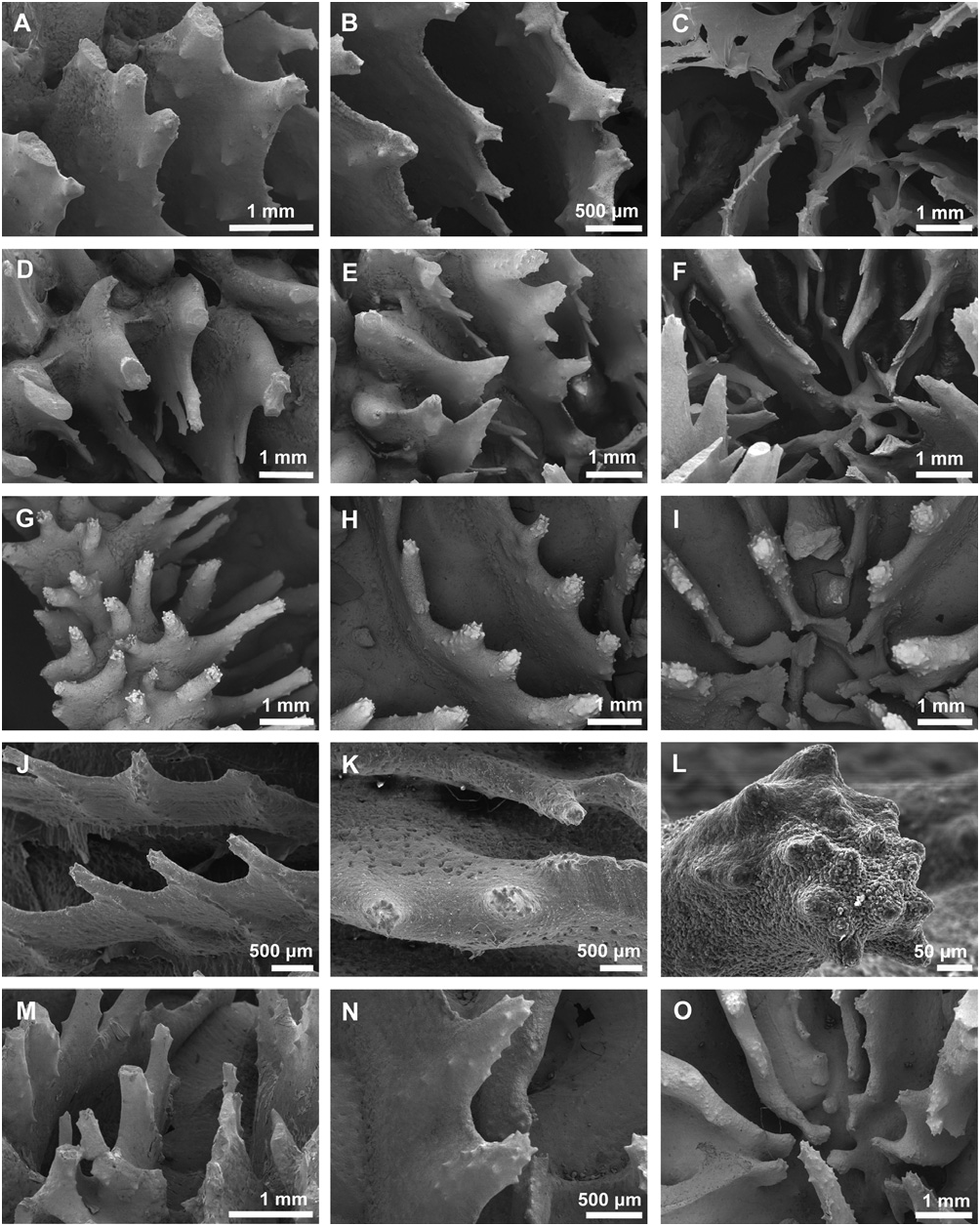
Micromorphology: high ( > 0.6 mm), widely spaced ( 1–2 mm), spine-shaped teeth, regularly arranged, with spiked tips resembling a Medieval spiked mace weapon; layered (banded) interarea of septal teeth; feeble or absent columella; paliform lobes; fine granules along the sides of teeth ( Fig. 19G–O View Figure 19 ).
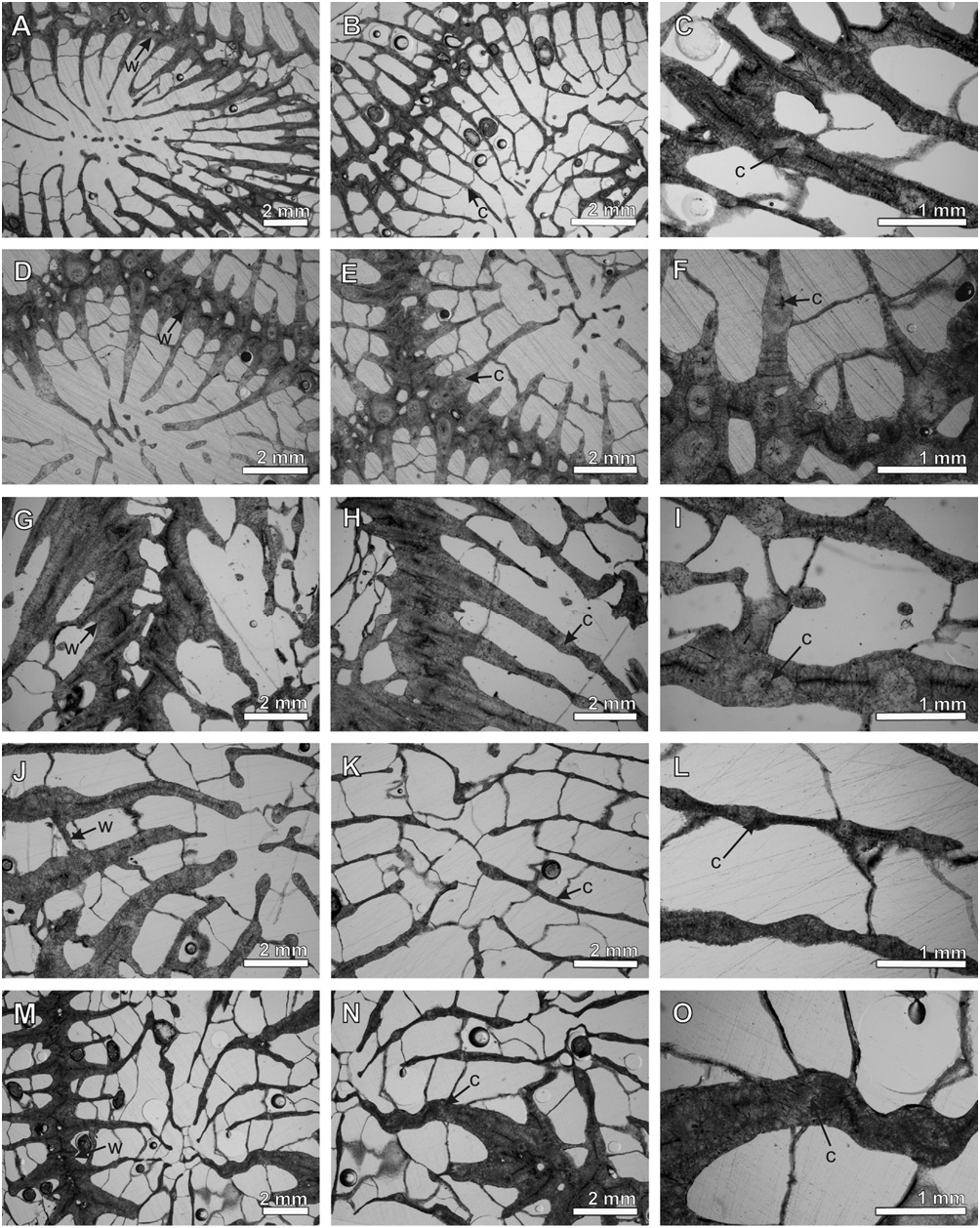
Microstructure: parathecal wall with trabeculothecal elements. Well-developed clusters of calcification centres encircled by concentric rings of fibrous thickening deposits; clusters cross medial lines; reduced thickening deposits ( Fig. 23G–O View Figure 23 ).
Included species: Mycetophyllia lamarckiana Milne Edwards & Haime, 1849: 258 [ holotype = MNHN- Scle910, Fig. 9E, F View Figure 9 , unknown locality].
Mycetophyllia aliciae Wells, 1973: 41 View in CoL , figs 25–28 [ holotype = USNM53496 About USNM , Fig. 14C, D View Figure 14 , Discovery Bay , Jamaica].
Mycetophyllia danaana View in CoL Milne Edwards & Haime, 1849: 259 [ holotype is from an unknown locality, and is lost ( A Andouche, pers. comm., 2007). We herein designate specimen SUI102772 View Materials ( FA1003 ) ( Fig. 14E, F View Figure 14 ) collected by H. Fukami at Bocas del Toro, Panama as the neotype.] .
Mycetophyllia ferox Wells, 1973: 40 View in CoL , figs 22–24 [ holotype = USNM53494 About USNM , Fig. 14G, H View Figure 14 , Eaton Hall , Jamaica].
Mycetophyllia reesi Wells, 1973: 36 View in CoL , figs 19–21 [ holotype = USNM53493 About USNM , Fig. 14I, J View Figure 14 , West Bull , Jamaica]
Remarks: In the classification system of Vaughan & Wells (1943) and Wells (1956), the genus Mycetophyllia is distinguished by having a meandroid colony form, long series, centres with lamellar linkage, discontinuous collines enclosing one or more continuous series, and a reduced or absent columella; colonies form initially by circumoral budding ( Vaughan & Wells, 1943: 193, 196). Our observations indicate that it also has paliform lobes. Mycetophyllia is generally similar to Isophyllia in microstructure and micromorphology. Isophyllia differs from Mycetophyllia in that Mycetophyllia lacks a coenosteum, and has confluent costosepta, centres with lamellar linkage, and paliform lobes, as described above.
Following Wells (1973), Cairns, Hoeksema & Land (1999) and Veron (2000) recognized five species of Mycetophyllia , whereas Zlatarski & Estalella (1982) recognized two ( Mycetophyllia lamarckiana , Mycetophyllia ? reesi ), the latter one of which was only questionably assigned to Mycetophyllia . Lang’s (1973) experimental studies of aggressive interactions found that Mycetophyllia ferox was the most aggressively superior, followed by Mycetophyllia reesi , but there was no interaction amongst Mycetophyllia lamarckiana , Mycetophyllia aliciae , and Mycetophyllia danaana . The five species of Mycetophyllia are distinguished primarily on the basis of colony form, which may be highly variable within species. Mycetophyllia reesi is distinguished by circumoral budding and no collines ( Fig. 14I, J View Figure 14 ); and Mycetophyllia ferox by intracalicular budding and narrow (~ 10 mm), meandering, sometimes discontinuous valleys with continuous forked collines ( Fig. 14G, H View Figure 14 ). Mycetophyllia aliciae , Mycetophyllia danaana , and Mycetophyllia lamarckiana all initially have circumoral budding (less pronounced in Mycetophyllia danaana ), followed by intracalicular budding, and straight, continuous valleys. The valleys in Mycetophyllia aliciae are multiserial and the collines discontinuous ( Fig. 14C, D View Figure 14 ), whereas the valleys in Mycetophyllia danaana and Mycetophyllia lamarckiana are uniserial. Mycetophyllia danaana and Mycetophyllia lamarckiana also tend to have less pronounced paliform lobes than the other three species (J. Lang, pers. comm.). Mycetophyllia danaana has deep valleys ( 10–12 mm), sometimes forming monticules (superficial ‘hydnophoroid pillars’ sensu Wells, 1973), discontinuous collines, and more septa (12–16 per cm; Fig. 14E, F View Figure 14 ); whereas Mycetophyllia lamarckiana has broad, shallow ( < 10 mm) valleys, continuous collines, and fewer septa (six to seven per cm; Fig. 14A, B View Figure 14 ). Danaher (1998) interpreted Mycetophyllia lamarckiana and Mycetophyllia danaana as being synonymous based on analyses of skeletal variation caused by environmental gradients related to energy and nutrient acquisition (functional plasticity).
GENUS SCOLYMIA HAIME, 1852: 279 View in CoL ( FIGS 9G, H View Figure 9 , 12A, B View Figure 12 , 18A–C View Figure 18 , 22A–C View Figure 22 )
Synonyms: Lithophyllia View in CoL Milne Edwards, 1857: 290 [ type species = Madrepora lacera Pallas, 1766 , by subsequent designation ( Felix, 1925: 100)].
Type species: Madrepora lacera Pallas, 1766: 298 ; by subsequent designation, Vaughan, 1901: 6. Holotype is lost. We herein designate specimen YPM 9036 collected by J. C. Lang in 1968 at Rio Bueno, Jamaica, as the neotype.
Original type species locality: ‘Mare Americanum’ ( Pallas, 1766: 298) [Recent].
Original descriptions:
1. ‘Je propose donc de laisser le nom de Caryophyllia à tous les polypiers qui présentent les caractères reconnus en 1828 par M. Stokes, et que M. Ehrenberg et nous-mêmes avons décrits depuis celui de Cyathina , en assígnant au genre Caryophyllia , Milne Edw. et J. Haime (Compt. rend. de l’Ac. des sc., t. XXVII: 491, 1848 – non Stokes), si tant est qui cette division mérite d’être conservée la dénomination de Scolymia que lui donne M. Jourdan dans la collection du Musée de Lyon. Cette restauration est d’autant plus importante, qu’il n’y a pour ainsi dire pas deux auteurs qui aient attribué la même un terme à cette regrettable confusion.’ ( Haime, 1852: 279)
2. ‘Coral cylindro-turbinate, cylindrical, or almost prismatical, in all ages attached to the ground by an expanded base, without a distinct epitheca. Costae prominent, roughly spinose, the uppermost spines being the strongest. Calicle shallow, circular, rarely oblong, rectangular or lobate. Septa of first and second cycles with their free edges lacerodentate, the teeth increasing in size from within outwards. Columella oblong in outline, consisting of thin trabeculae, its surface finely papillose or imbricate.’ ( Brüggemann, 1877: 301)
Subsequent morphological descriptions ( Matthai, 1928, and later): = Mussa in Matthai (1928: 202–208); = Mussa in Vaughan & Wells (1943: 195); = Mussa in Wells (1956: F418); Wells (1964: 375–384); Laborel (1969: 217–222); Lang (1971: 952–959); Wells (1971: 960–962); Walton Smith (1971: 92); Zlatarski & Estalella (1982: 157–165); Veron (2000: vol. 3: 66–71); Neves et al. (2006: 45–54).
Diagnosis: Macromorphology: solitary, with rare intracalicular budding; large calices ( > 4 cm), more than four septal cycles, unequal; well-developed, spongy ( > three threads) columella with lamellar linkage; reduced epitheca; no septal or paliform lobes; abundant endotheca ( Figs 9G–H View Figure 9 , 12C–H View Figure 12 ).
Micromorphology: high ( > 0.6 mm), widely spaced ( 1–2 mm), spine-shaped, pointed teeth, regularly arranged; smooth interarea of septal teeth; teeth in major and minor septal cycles differ in size; spongy columella, with columellar teeth differing in size and shape from septal teeth; spiky, aligned granules ( Fig. 18D–L View Figure 18 ).
Microstructure: parathecal wall with trabeculothecal elements. Widely separated ( > 1.2 mm), welldeveloped clusters of calcification centres encircled by concentric rings of fibrous thickening deposits; clusters cross weak medial lines; moderate thickening deposits ( Fig. 22D–L View Figure 22 ).
Included species: Scolymia lacera ( Pallas, 1766: 298) [ holotype is from ‘Mare Americanum’, and is lost; neotype (herein designated) = YPM9036 About YPM , Fig. 9G, H View Figure 9 , Rio Bueno , Jamaica] .
Scolymia cubensis ( Milne Edwards & Haime, 1849: 238) [ holotype is from Cuba, and is lost ( A. Andouche, pers. comm., 2009); neotype (herein designated) = YPM7569 About YPM , Fig. 12E, F View Figure 12 , Runaway Bay , Jamaica] .
Scolymia wellsi View in CoL ( Laborel, 1967: 107, figs 1–3) ( holotype = MNHN-Scle20175, Fig. 12G, H View Figure 12 , Abrolhos Archipelago , Brazil) .
Remarks: Following Zlatarski & Estalella (1982), Veron (2000) synonymized Scolymia lacera (Pallas) with Mussa angulosa (Pallas) , and redesignated Scolymia cubensis ( Milne Edwards & Haime, 1849) as the type species of Scolymia . Our observations indicate that Scolymia lacera and Mussa angulosa are distinct. Scolymia differs from Mussa in its colony form, more extensive thickening deposits, more closely spaced clusters of calcification centres ( < 1.8 mm), smooth interarea of septal teeth, and distinctive paddle-shaped columellar teeth. When budding, it also has lamellar linkage, in contrast to trabecular linkage in Mussa . We therefore retain Madrepora lacera Pallas, 1766 , as the type species of Scolymia .
As described by Wells (1964: 375–376), the genus Scolymia is characterized by wholly dentate septal margins, dentations four to six (per) cm on larger septa, large calices ( > 4 cm), centres with trabecular linkage, and sparse septal granules. Scolymia is distinguished from the genus Homophyllia [ type species = Homophyllia australis ( Milne Edwards & Haime, 1849)] on the basis of number of dentations [ten to 12 (per) cm in Homophyllia ] and calice size ( < 4 cm in Homophyllia ). Scolymia is distinguished from the genus Parascolymia [ type species = Parascolymia vitiensis ( Brüggemann, 1877) ] on the basis of septal granulation (more numerous and thicker in Parascolymia ). Wells (1964) also indicates that Scolymia has trabecular linkage amongst centres, as opposed to Parascolymia . However, our observations show that linkage in Scolymia is indeed lamellar. Veron (2000: vol. 3: 66–71) later assigned Homophyllia australis and Parascolymia vitiensis to the genus Scolymia , presumably because of their monocentric colony form, and their ‘large, regular, blunt teeth’. Following the molecular results of Fukami et al. (2004), we restrict the definition of the genus to include only Atlantic taxa. The genus, therefore, does not include the following species described in Veron (2000): Scolymia australis ( Milne Edwards & Haime, 1849: 310) [= Homophyllia Brüggemann, 1877 ]; Scolymia vitiensis Brüggemann, 1877: 304–305 [= Parascolymia Wells, 1964 ].
Wells (1971) recognized two species of Scolymia ( Scolymia lacera , Scolymia cubensis ), which were supported by Lang’s (1971) experiments on aggressive interactions. Laborel (1967) described a third species, Scolymia wellsi from Brazil. Although Zlatarski & Estalella (1982) synonymized the three species, Cairns, Hoeksema & Land (1999) and Neves et al. (2006) recognized them as being distinct. As described by Neves et al. (2006), the three species can be distinguished on the basis of septal dentation. Scolymia wellsi has irregular teeth that are sometimes fused forming porous septa ( Fig. 12G, H View Figure 12 ); Scolymia cubensis has long and slim teeth ( > five per cm) that are awl-shaped ( Fig. 12E, F View Figure 12 ); Scolymia lacera has large and stout teeth ( < five per cm) that are subtriangular ( Fig. 12C, D View Figure 12 ). Wells (1971) and Lang (1971) further noted that differences between lower and higher cycle septa are more pronounced in Scolymia lacera , its calices are more concave, and its maximum calice diameters are larger ( 15 cm as opposed to 10 cm in Scolymia cubensis ). The corallum of Scolymia wellsi is similar to Scolymia cubensis , although even smaller ( < 6 cm). Given the morphological similarities amongst the three species, we have designated neotypes for Scolymia lacera and Scolymia cubensis using material collected and studied by Lang (1971).





| YPM |
Peabody Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Mycetophyllia
| Budd, Ann F., Fukami, Hironobu, Smith, Nathan D. & Knowlton, Nancy 2012 |
Mycetophyllia aliciae
| Wells JW 1973: 41 |
Mycetophyllia ferox
| Wells JW 1973: 40 |
Mycetophyllia reesi
| Wells JW 1973: 36 |
Scolymia wellsi
| Laborel J 1967: 107 |
SCOLYMIA HAIME, 1852: 279
| Haime J 1852: 279 |
Mycetophyllia danaana
| Milne Edwards H & Haime J 1849: 259 |
MYCETOPHYLLIA MILNE EDWARDS & HAIME, 1848: 491–492
| Milne Edwards H & Haime J 1848: 492 |