
Lates mariae, Steindachner, 1909
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2004.00111.x |
|
publication LSID |
lsid:zoobank.org:pub:242EC75C-52E2-468A-ABD5-C092D7F3908E |
|
persistent identifier |
https://treatment.plazi.org/id/AE3187EB-2F23-FFDA-2A09-E378943FFE32 |
|
treatment provided by |
Diego |
|
scientific name |
Lates mariae |
| status |
|
G Lates mariae View in CoL frontals and possibly of the supraoccipital. The supraoccipital limit observed in † L. bispinosus is similar to L. niloticus ; that indicates the presence of a pleurosphenoid pedicle in the fossil species. A saggital crest develops along the whole supraoccipital length (see below). In L. stappersi , the supraoccipital lateral flanges are confined anteriorly whereas they develop all along the crest in other latids.
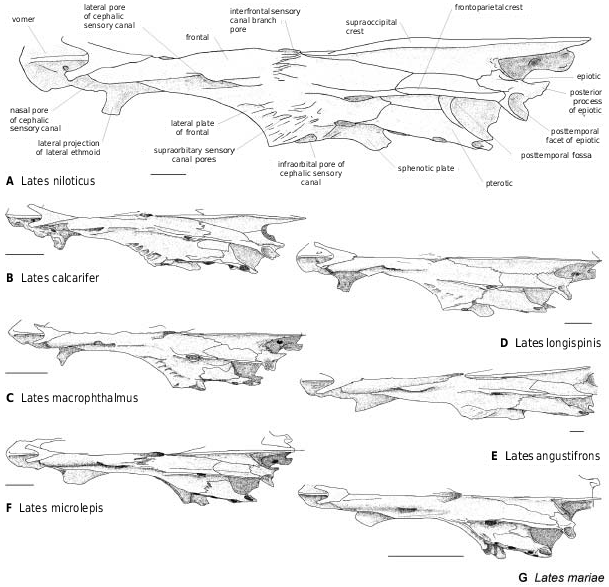
The frontal lateral plate lies in front of the sphenotic ( Figs 7–11 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 ). Its development beyond the orbit seems to be correlated with that of the pleurosphenoid pedicle: it is absent in Psammoperca ( Fig. 11D View Figure 11 ), reduced in L. mariae ( Figs 8G View Figure 8 , 10G View Figure 10 ) and L. stappersi ( Fig. 8H View Figure 8 ), little developed in L. macrophthalmus ( Figs 8D View Figure 8 , 10D View Figure 10 ) and L. longispinis ( Figs 8C View Figure 8 , 10C View Figure 10 , 11C View Figure 11 ), and well developed in L. niloticus ( Figs 7 View Figure 7 , 8A View Figure 8 , 10A View Figure 10 , 11B View Figure 11 ), L. calcarifer ( Figs 8B View Figure 8 , 10B View Figure 10 ) and L. angustifrons ( Figs 8E View Figure 8 , 10E View Figure 10 , 11A View Figure 11 ), particularly in larger individuals. Concerning the other latid species, it is well developed in † L. bispinosus ( Fig. 9 View Figure 9 ) and reduced in † E. gracilis ; these indicate, respectively, the presence and the absence of a pleurosphenoid bridge. The supraorbital sensory canal openings lie on the dorsal face of the plate ( Fig. 11A–C View Figure 11 ), or in the edge of the bone when the plate is reduced or absent ( Fig. 11D View Figure 11 ).
The latid epiotic ( Figs 9 View Figure 9 , 10 View Figure 10 ) has a well-developed facet for the articulation of the upper limb of the posttemporal that projects latero-posteriorly, a strong horizontal posterior process that projects medial to the articular facet, and the anterior bor- der inserts between the supraoccipital and the parietal. The posterior process varies in length and shape within a single species and sometimes between both left and right epiotics of a single individual (bifurcated in some individuals). In addition, the observation of several specimens of Lates niloticus and L. calcarifer supports a probable sizecorrelated variation, and species with a narrow neurocranium have a thinner posterior process and a smaller articular facet.
Both Recent and fossil latids bear fronto-parietal crests lateral to the supraoccipital crest ( Figs 8–10 View Figure 8 View Figure 9 View Figure 10 ; in † Eolates aquensis specimens only the saggital zone is suitable for observations). The height of the crests is inversely correlated with the neurocranial width. Van Neer & Gayet (1988) and Greenwood (1976), respectively, observed this correlation within a species ( Lates niloticus ) and among the Tanganyikan species. The crests seem to develop more in smaller individuals.
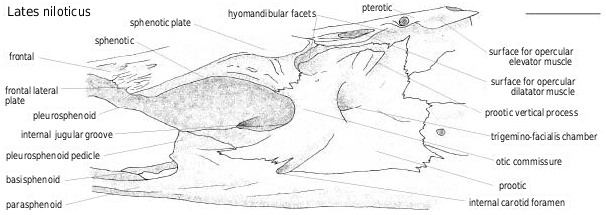
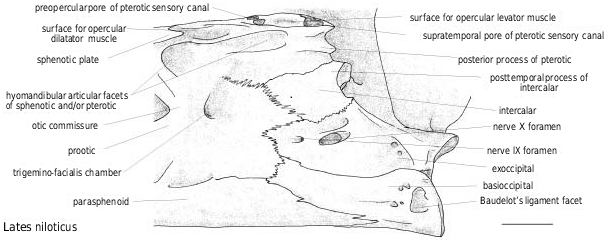
On the pterotic ( Figs 9 View Figure 9 , 12 View Figure 12 ), the oval-shaped surface for the opercular levator muscle is placed between the level of the two posterior openings of the cephalic sensory canal. The pointed posterior process develops under the supratemporal posterior facet of the pterotic. In some specimens, it develops back toward the posttemporal facet of the intercalar but in other specimens it does not even reach the anterior part of the intercalar. This is also the case for the posttemporal facet of the pterotic. The dimensions and directions of the processes and facets are variable and do not correlate with the neurocranial dimensions.
The cephalic sensory canal ( Figs 9–11 View Figure 9 View Figure 10 View Figure 11 ) runs in a bony tube with several openings. The main openings are constant in latid fish. On the nasal, the canal runs in a tube with an anterior opening for the sensory branch to the first infraorbital, and a posterior one toward the frontal. On the frontal, the lateral opening is placed at the level of the posterior tip of the lateral ethmoid [unknown in † Eolates ]; posteriorly, the interfrontal branch opens between the frontals in front of the anterior tip of the supraoccipital (e.g. in † Lates bispinosus , Fig. 9 View Figure 9 ), and on the lateral side of the frontal, the pore for the sensory branch to both the sphenotic and the pterotic is placed at the intersection of the sphenotic, pterotic and frontal. On the dermosphenotic, the canal runs in a tube with an opening for the branch to the infraorbital series. On the pterotic itself, two main pores allow the canal to reach the preoperculum (posterolateral opening) and the supratemporal (posterodorsal opening). The pores are smaller relative to the size of the skull in larger individuals. There are supernumerary openings on the pterotic; their count varies depending on the species and the individuals and is largest in Psammoperca ( Fig. 11D View Figure 11 ).
The occipital region is poorly known in fossils. The walls of the braincase are flat in Recent latids other than Psammoperca ( Fig. 13A View Figure 13 ), Lates macrophthalmus , L. microlepis , and larger L. mariae (which had the walls inflated in a specimen with a 25-mm-long neurocranium). In posterior view, the braincase of these species shows a basal inflation. Therefore, the inflation is observed in species with a curved parasphenoid, except in the case of small L. stappersi and large L. mariae (which have a curved parasphenoid but flat braincase walls).
The intercalar ( Figs 9 View Figure 9 , 12 View Figure 12 ) has the posttemporal articular facet placed on a bulky knob.
The exoccipitals ( Figs 12 View Figure 12 , 13 View Figure 13 ) articulate with the first centrum by facets that join above the basioccipital facet. In Lates ( Fig. 13B View Figure 13 ), the facets are beanshaped and participate in forming the base and part of the walls of the foramen magnum, whereas in Psammoperca ( Fig. 13A View Figure 13 ), they are teardrop-shaped and only form the base of the foramen magnum. Moreover, as in all the other percoids examined, the large foramen for nerve X (vagus) and the small and anteriormost foramen for nerve IX (glossopharyngial) are broadly separated.
The basioccipital ( Fig. 12 View Figure 12 ) usually bears relatively smaller surfaces for the Baudelot’s ligament in larger individuals. They are pointed or rounded in shape, and flatter in Tanganyikan species. Anterior to these are one or two depressions of variable size and more or less ornamented with small pores. As for the vomer, the suture of the basioccipital with the parasphenoid has interdigitations that seem shorter in larger individuals, as described by Van Neer & Gayet (1988), in L. niloticus . The median ventral groove on the basioccipital never reaches the parasphenoid, in L. niloticus ( Van Neer & Gayet, 1988) , nor in any other Recent latid (unknown in fossil species).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |