
Tatia
|
publication ID |
https://doi.org/ 10.1590/S1679-62252008000300022 |
|
persistent identifier |
https://treatment.plazi.org/id/AD092F4C-FFC0-FF82-FEBC-1342A3225D86 |
|
treatment provided by |
Carolina |
|
scientific name |
Tatia |
| status |
|
Tatia View in CoL View at ENA Miranda Ribeiro
Tatia View in CoL Miranda Ribeiro, 1911: 360 [type species: Centromochlus intermedius Steindachner, 1877 View in CoL , by subsequent designation by Jordan, 1920: 545. Gender: feminine].
Diagnosis. Tatia is distinguished among the Centromochlinae by three uniquely derived features. The hyomandibula is elongate anterodorsally, not contacting the narrow metapterygoid and, instead, connected only to the trapezoidal quadrate ( Fig. 2 View Fig ); anal-fin base of adult males is reduced (anal-fin base length 3.3-8.0% SL); and caudal peduncle is compressed and deep (caudal-peduncle depth 10.1-18.6% SL), with a middorsal keel posterior to adipose fin.
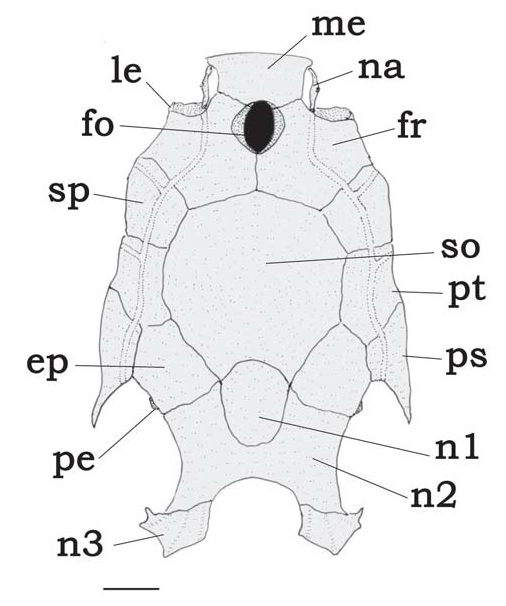
A unique combination of restricted characters aids in distinguishing Tatia : first (anteriormost) nuchal plate present ( Fig. 3 View Fig , n 1 View Fig ); infraorbital 1 bone short, limited to anterior corner of orbit; eye moderately large, between 17.4-42.9% HL; maxilla shorter than or same length as autopalatine; retractor tentaculi muscle absent, functionally substituted by maxillomandibular ligament (Sarmento-Soares & Porto, 2006); adult males with modified anal fin (all species), and first unbranched anal-fin ray non-segmented or with segments fused (except in T. brunnea ).
Generic description. Tatia comprises species with somewhat robust head; caudal peduncle much compressed laterally and deep; well-developed adipose eye lid; eye moderately large,
and latero-dorsally located. Following characters used to distinguish species of Tatia (illustrations based on T. gyrina ): Head: cranial fontanel always present ( Fig. 3 View Fig , fo); premaxilla transversely elongated, meeting its counterpart medially; autopalatine tubular, oriented obliquely to longitudinal axis of body; maxilla very small, shorter than or same length as autopalatine; three nuchal plates present ( Fig. 3 View Fig , n 1 View Fig , n 2 View Fig , n 3 View Fig ). Epioccipital process small, connected to third nuchal plate by ligaments ( Fig. 3 View Fig , pe). Mullerian ramus with distal tip shaped as slightly curved disc, its concave face superficially adjoined to anterior wall of gasbladder. Prevomer usually with well developed arrow-shaped lateral processes or with short lateral process. Prevomerian teeth present in T. galaxias and T. intermedia .
Suspensorium ( Fig. 2 View Fig ): Suspensorium bones in Tatia similar among all species and distinctive among Centromochlinae . Hyomandibula narrow, elongate, projected anteriorly as short membranous lamina ( Fig. 2 View Fig , hy). Anterior laminar projection of hyomandibula sutured only to quadrate, through cartilage ventrally and deeply dentate suture dorsally. Metapterygoid conical, dorsally sharp, with little laminar extension ( Fig. 2 View Fig , mt), joined to quadrate via cartilage block only ( T. gyrina , T. caxiuanensis , T. meesi , T. strigata ) or cartilage plus dentate suture ( T. aulopygia , T. boemia , T. brunnea , T. dunni , T. galaxias , T. intermedia , T. neivai , T. nigra ). Suspensorium deeply notched anterodorsally, thus metapterygoid not contacting hyomandibula. Quadrate trapezoidal, with broad base ( Fig. 2 View Fig , qu), connected to preopercle, hyomandibula and metapterygoid; long preopercle ( Fig. 2 View Fig , po) ventral margins sutured to both quadrate and hyomandibula; suprapreopercle present as short or robust canal bone ( Fig. 2 View Fig , sp); opercle laminate and broadly subtriangular ( Fig. 2 View Fig , op).
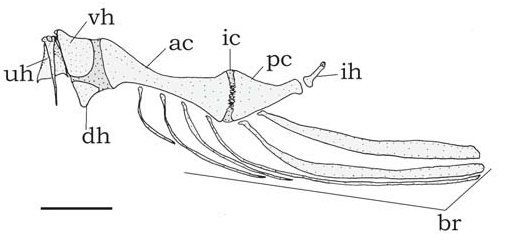
Hyoid arch ( Fig. 4 View Fig ): Urohyal narrow ( Fig. 4 View Fig , uh); short dorsal hypohyal ( Fig. 4 View Fig , dh) associated with comparatively large ventral hypohyal ( Fig. 4 View Fig , vh); anterior ceratohyal well developed ( Fig. 4 View Fig , ac), posterior ceratohyal smaller ( Fig. 4 View Fig , pc); branchiostegal ray articulated to hyoid arch; branchiostegal rays 5-7; 3-5 on anterior ceratohyal and two on posterior ceratohyal; posteriormost usually largest and flattened. No branchiostegal ray associated with interceratohyal cartilage between bones.
Pharyngeal (gill) arches ( Fig. 5 View Fig ): Basibranchial 1, in contact with urohyal ( Fig. 5 View Fig , ab), anterior to basibranchial 2; basibranchial 2 forming osseous rod, broadest anteriorly ( Fig. 5 View Fig , bb2) and usually separated by gap from basibranchial 3; basibranchial 3 shorter ( Fig. 5 View Fig , bb3), also broadest anteriorly; basibranchial 4 large, flattened and completely cartilaginous ( Fig. 5 View Fig , bb4); basibranchial 2 bordered laterally by cartilaginous head of hypobranchial 1; basibranchial 3 between cartilaginous head of hypobranchial 2 and cartilaginous hypobranchial 3; basibranchial 4 bordered laterally by cartilaginous head of ceratobranchial 4 and caudally by cartilaginous head of ceratobranchial 5. Hypobranchial 1 mostly osseous, elongate and expanded laterally, subtriangular, with cartilaginous ends; hypobranchial 2 mostly cartilaginous, subtriangular, with medial osseous part; hypobranchial 3 completely cartilaginous, more trapezoidal ( Fig. 5 View Fig , hb 1-3); hypobranchial 4 absent. Five ceratobranchials present, mostly ossified, with cartilage on both ends ( Fig. 5 View Fig , cb 1-5). First ceratobranchial supporting single row of rakers (not illustrated); fifth ceratobranchial expanded postero-medially to support lower pharyngeal toothplate with short conical teeth. Five epibranchials, all but fifth largely ossified except for cartilaginous ends; epibranchials 1 and 2 rod-like, without rakers; epibranchial 3 with posterior uncinate process; epibranchial 4 broad, with laminar extension; epibranchial 5 cartilaginous, reduced, located in axil between cartilaginous ends of epibranchial 4 and ceratobranchial 4. Pharyngobranchial 1 absent; pharyngobranchial 2 short, cartilaginous, somewhat rounded, placed between anteromedial cartilaginous tips of epibranchials 1 and 2; pharyngobranchial 3 elongate, ossified, with expanded posterior border; pharyngobranchial 4 ossified, supporting upper pharyngeal tooth plate with short conical teeth.
Lateral line system: Infraorbital 1 short, with ventro-lateral process either clearly restricted to anterior border of eye or almost reaching ventral border of eye; infraorbitals thin and canalicular, forming incomplete or complete infraorbital series. Lateral line on body straight, inconspicuous, with ossified canal bones just anteriorly.
Fins: Dorsal fin with major spine preceded by small dorsal locking spine and followed by 3-5 branched rays; pectoral fin with one spine plus 3-5 branched rays; pelvic fin with one unbranched plus 5 branched rays, margin rounded; adipose fin above anal-fin base; anal fin with 3 unbranched plus 6-8 branched rays; caudal fin with 8+9 principal rays, forked, lobes with rounded tips; dorsal caudal-fin lobe slightly elongate in males of some species.
Ribs and vertebrae: Pleural ribs 7-11; first rib thicker than others; ribs becoming progressively smaller posteriorly. Post- Weberian vertebrae 30-39.
Sexual dimorphism. In catfishes of subfamily Centromochlinae the anal fin of sexually mature males is modified into a sperm conductor with the anal-fin rays and proximal radials directed posteriorly, aligned nearly parallel to vertebral axis ( Ferraris, 1988; Soares-Porto, 1998). In mature males the anal fin is strongly modified such that it is supported by enlarged and joined proximal anal-fin radials ( Ferraris, 1988; Soares-Porto, 1998) and by partially ossified distal radials. Anal-fin proximal radials are associated with the thickened and lengthened hemal spines of adjacent vertebrae. Modified male anal fin has three developed unbranched rays. The first unbranched ray is shortest, about one-quarter length of second unbranched ray, and with segments fused (except in T. brunnea ). Some species have the first unbranched anal-fin ray preceded immediately by a tegumentary keel. The second and third unbranched rays are thick. In the Centromochlinae , Tatia is the only genus with species that bear segments that curve outwards, forming antrorse denticulations along the anterior margin of the third unbranched anal fin ray in adult males.
Remarks. Two of the nominal species assigned to Tatia by Ferraris (2007) are not herein recognized as members of this genus. Both Tatia musaica Royero and Tatia simplex Mees lack the derived characters listed above for Tatia : thin, anterodorsally elongate hyomandibula (vs. broad in both C. musaicus and C. simplex ); caudal peduncle laterally compressed (vs. ellipsoid); proportionally deeper caudal peduncle, depth 10.1-18.6% SL (vs. 7.9-9.6% SL in C. musaicus and 9.2- 10.0% SL in C. simplex ); first nuchal plate present (vs. ab- 1989; Franke, 1990), but a reproductive behavior is not yet reported. During reproductive phase, the anal-fin rays change their position probably channeling for discharge of seminal ducts, acting as a sperm conductor during reproduction. Such a morphological change was observed in some males of Centromochlus romani and also in T. nigra . The sperm probably runs along the spoon shaped anal-fin rays, and the fish may press the fin against the belly of the female, for inseminating. Morphological evidences suggesting inseminating reproductive mode in Tatia are the intumescent female genital papilla (as in T. gyrina and T. nigra ) and the spoon-shaped laterally curved male anal fin (observed in T. nigra ).A detailed analysis of morphological changes of the reproductive tract needs to be accomplished by histological preparations, and is out of the scope of present contribution.
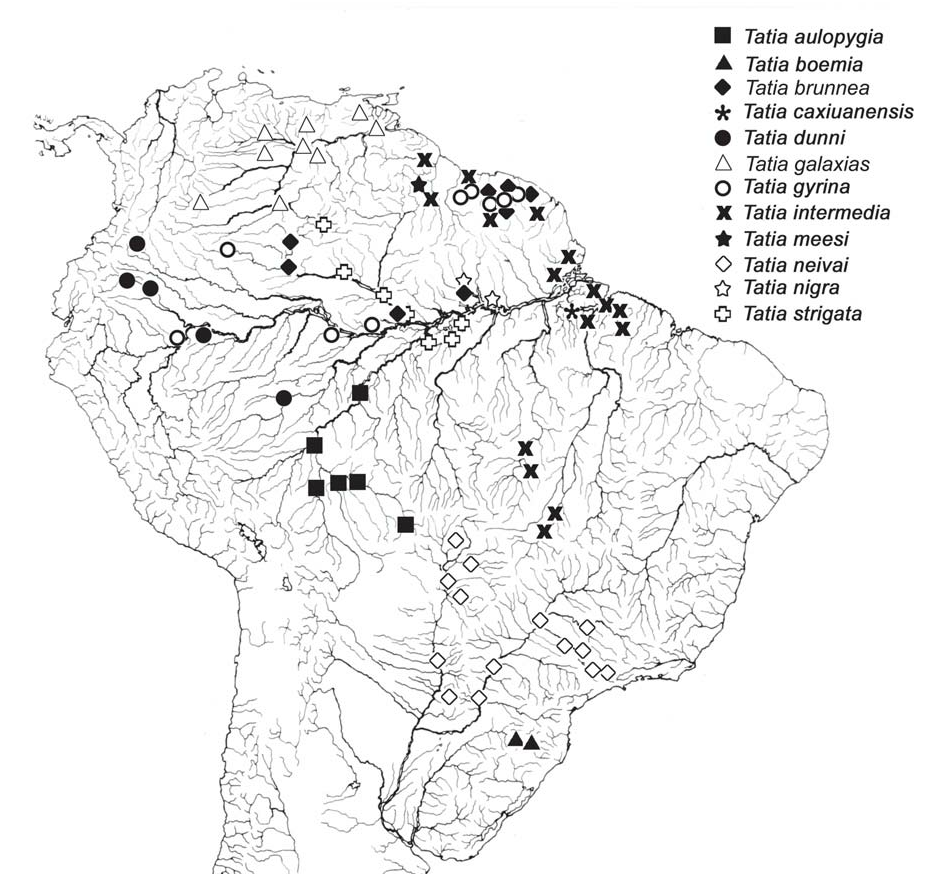
Distribution. Species of Tatia are broadly distributed in the regions east of the Andean Cordilleras ( Fig. 6 View Fig ). Tatia ranges from the Guarapiche river, Orinoco basin, in Venezuela, to the Pelotas river, Uruguay drainage, in Brazil. The group is found in major continental river basins, such as the Amazon, Orinoco and Paraná-Paraguay; as well as in the Essequibo and small coastal drainages in French Guiana, Suriname and Amapá and Marajó island in northern Brazil. Although well distributed in cis-Andean South America, the group is not known from certain drainage basins, including the São Francisco and small coastal river drainages of northeastern and far eastern Brazil. Tatia is found, however, in headwaters of Paraíba do Sul basin, São Paulo State. The absence of Tatia from certain areas may be a function of inadequate collections from those river basins or may reflect actual disjunct distributions of the group as whole.
sent); maxilla shorter than or same length as autopalatine (vs. longer than autopalatine). These two species are incertae sedis in Centromochlinae (see Table 1), based in part on interrelationships analysis by Soares-Porto (1996), and are herein provisionally allocated under Centromochlus .
The monophyly and interrelationships among species of Tatia were hypothesized by Soares-Porto (1998), who considered the clade T. gyrina plus T. creutzbergi as sister to all other Tatia species.
Inseminating reproductive mode (sensu Burns et al., 1997) has been documented within theAuchenipteridae (von Ihering, 1937; Loir et al., 1989; Meisner et al., 2000).In the centromochlin catfishes, inseminating is supposed to take place (Loir et al.,



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Tatia
| Sarmento-Soares, Luisa Maria & Martins-Pinheiro, Ronaldo Fernando 2008 |
Tatia
| Jordan, D 1920: 545 |
| Miranda Ribeiro, A 1911: 360 |