
Eumasia thomasii, Unnikrishnan & Sobczyk & Jose & Jose, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5200.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:27594EE8-4A77-4D6C-A411-D9710C7DF756 |
|
DOI |
https://doi.org/10.5281/zenodo.7262116 |
|
persistent identifier |
https://treatment.plazi.org/id/AB287B37-197B-E102-FF60-DA28FB0AFE33 |
|
treatment provided by |
Plazi |
|
scientific name |
Eumasia thomasii |
| status |
sp. nov. |
Eumasia thomasii View in CoL sp. nov.
( Figs 1–11 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 )
Type material. Holotype. ♂, INDIA, Kerala, Vattavada, Koviloor , 10.18211° N, 77.25923°, 11/07/2022, with larval case, leg. Usha A U. GoogleMaps Holotype will be deposited at the Zoological Survey of India Museum, Calicut, Kerala after submission of thesis of the first author and is at present at St. Thomas College , Thrissur .
Paratypes: 2 females (both with larval case), same data as holotype, 10/07/2022, 25 larval cases with pupal exuviae, leg. Usha A U.
Etymology. The adult female in most bagworm species is characterized by rudimentary appendages and eyes and no wings. But in this species both male and female are winged, have well developed eyes, legs, and antennae. Therefore; the male and female adult moths are almost twin like. The word Thomas means ‘twin’ in aramic. The institution “St. Thomas College” in which the major part of the research was carried out is named for St. Thomas. Hence the species name “ thomasii ” alludes to the similar appearance of males and females and is also a homage to the institution.
Description. Holotype ♂ ( Fig. 1 View FIGURE 1 ), wingspan 8–8.3 mm, body length 2.5–3 mm, forewing length 4.1 mm including fringes, width 1.1 mm, WI 3.7 mm.
Head. Vertex roughly covered with pale golden-brown hairs. Eyes large, inter ocular index, EI 1.2 mm.Antenna short, filiform, 1.7 mm long, and 25 segments ( Fig. 2a View FIGURE 2 ). Segments sparsely ciliated, especially ventrally, dorsally with a fan-shaped process of dark brown scales, angle about 35° (according to Hättenschwiler 1998).
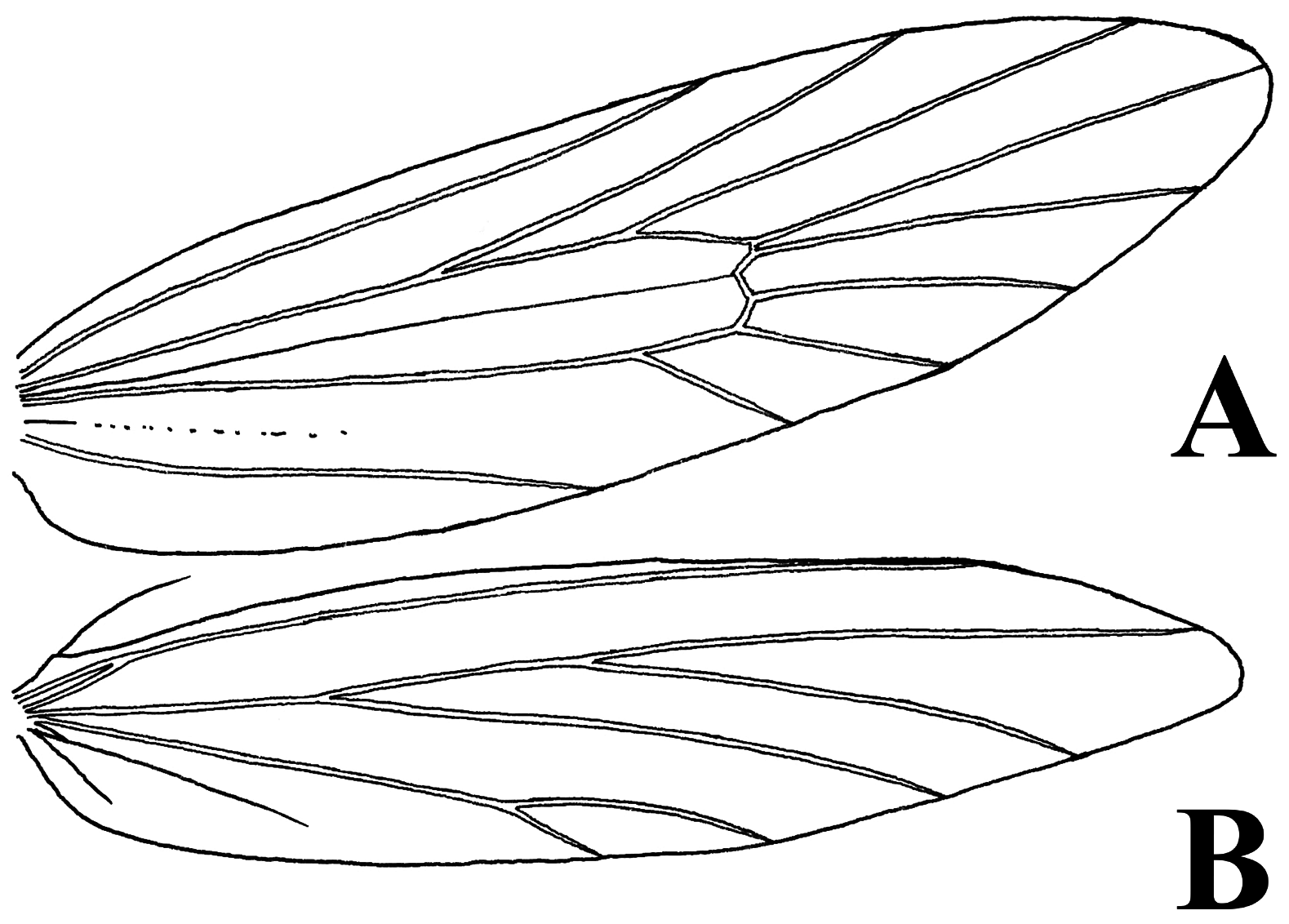
Thorax. Densely covered with long brown scales. Forewing ( Fig. 3a View FIGURE 3 ) narrow and lanceolate, costa straight and termen tapering at apex. Discoidal cell is half the length of the forewing. Median stem of dc complete but faded from middle to apex dc. Dc bordered externally with transverse veins (not open, as in most Eumasia species). Venation: forewing with nine veins, seven veins from the dc. Sc straight and ended up in the middle length of costa. R1 from dc is parallel to the vein Sc but originating different length from the base. R2+3 and R4+5 run parallelly and end up on the costal margin at different lengths. M1 and R4+5 originate apically from dc reaching to apex. Veins R4+5 and M1 are not stalked at base but originate from the same point on the apex of dc. Veins M2+3, CuA1 and CuA2 are short and parallel to each other.
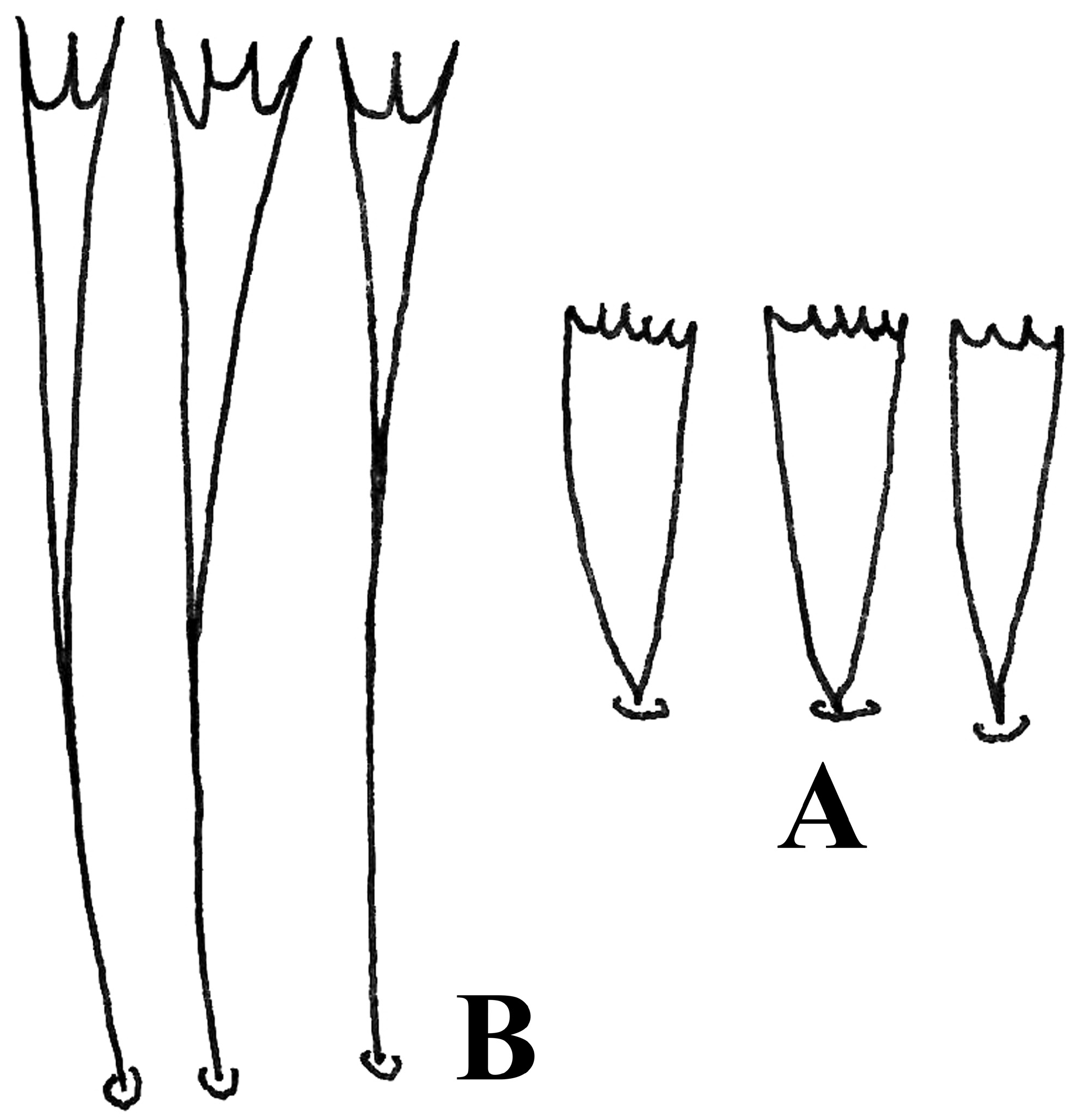
Hindwing ( Fig. 3b View FIGURE 3 ) is narrow and small, costa straight and vein Sc+R1 run adjacent to costa. Vein Rs and M1 are stalked and terminating to apex and termen respectively. M1 and M2+M3 run parallelly to termen. CuA1 and CuA2 are stalked and short. Anal veins are very short and almost invisible on the wings. Forewings thickly covered with broad, mostly 5–6 pointed scales ( Fig. 4a View FIGURE 4 ) (classes 5–6 after Sauter 1956). Ground colour of the upper side of the forewing is greyish white marked with blackish brown spots. Anterior margin without clear dark spots. An indistinct oblique band of dark scales from one-fourth of the anterior margin to half of the posterior margin.Another band from half anterior margin to half posterior margin. A third band slightly distal to it. A most conspicuous feature is a broad band of dark scales at three-quarters wing length. Distal to this, isolated dark scales towards the apex and outer margin. Fringes whitish, narrow, unicoloured, usually 3-pointed distally ( Fig. 4b View FIGURE 4 ).
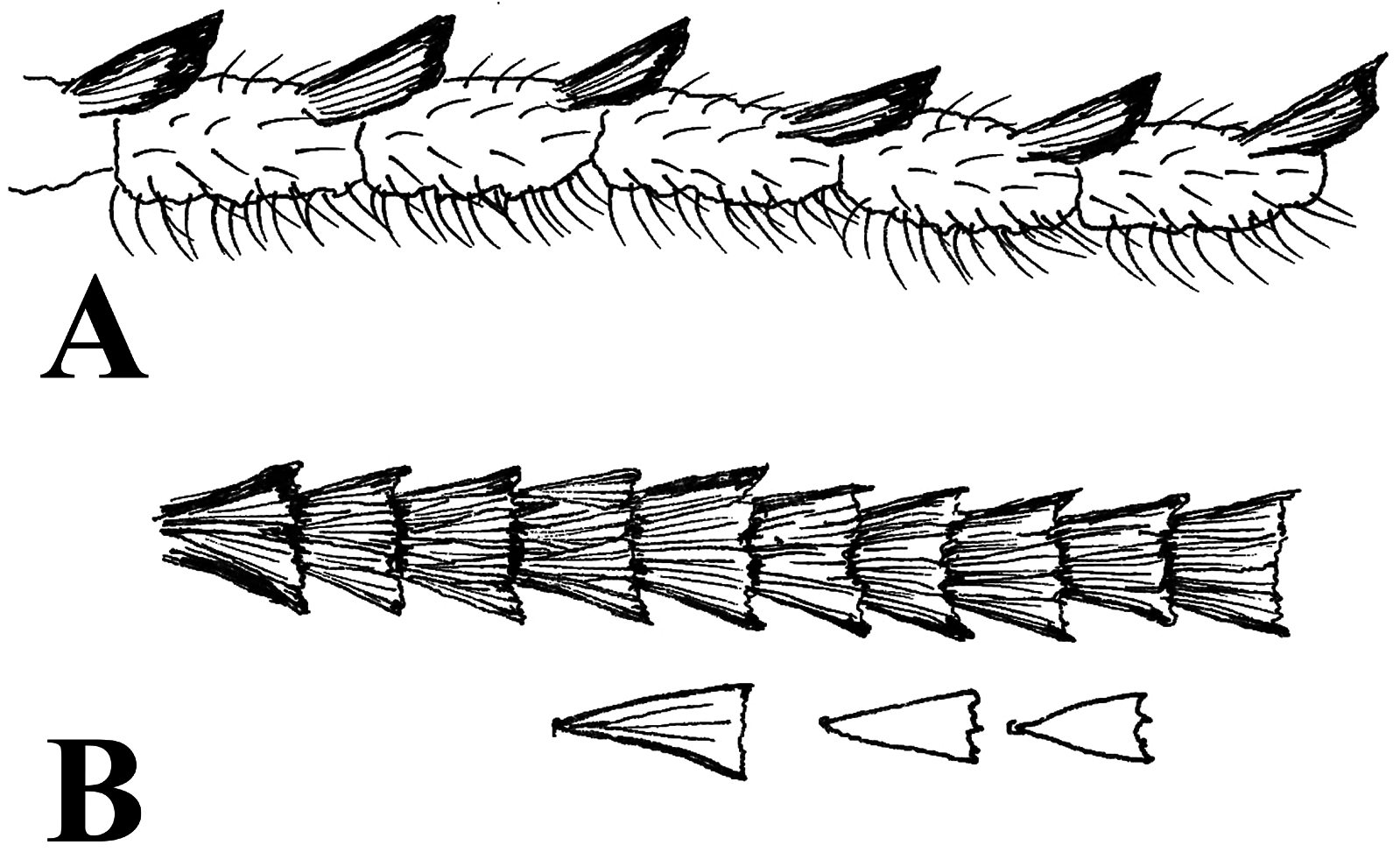
Hindwing ground colour silver white without any spots and markings, covered with white greyish coloured scales. Scales narrower as fore wing, more rounded distally, few indentations. Outer margin with long whitish fringes. Forelegs ( Fig. 5 View FIGURE 5 ) ventrally with long hairbrush along with the length of tibia, hind legs are very long, 4.5 mm in length, covered with long whitish hairs on tibia and shorter hairlike scales on tarsal segments.
Abdomen. Abdomen long, thickly covered with blackish brown scales.
Genitalia ( Fig. 6 View FIGURE 6 ). 0.86 mm long, Tegumen arched anteriorly and concave Valva elongate and symmetrical, presence of hairs anteriorly. Vinculum V shaped and saccus very long and pointed at apex. Saccus two-thirds the length of the genitalia. Phallus very long and straight tube-like structure, with pointed tip, length 0.52 mm.
Paratype ♀ ( Fig. 7 View FIGURE 7 ). Winged; Body 3.5–4.2 mm long, stout and larger than males; wingspan 8–8.8 mm including fringes, sizes of the wings of female and male are same, forewing length including fringes 4.1 mm, width 1.1 mm, forewing index 3.7 mm. Vertex is covered with thick pale golden coloured hairlike scales, antennae ( Fig. 2b View FIGURE 2 ) filiform, 23–25 segments, length 1.6 mm, completely covered with broad scales arranged in rings, slightly projecting distally, without cilia. Thorax is pale brown, sparsely covered with scales. Abdomen long, stout, pale-yellow, 3 mm long, white greyish scales on the abdominal segments and golden coloured hair tuft on the last abdominal segment. Forewing thickly covered with light brown and blackish scales; blackish scales are arranged as spots on the forewings. Hindwings are without markings and patterns, densely covered with white greyish scales, outer margins of both the wings are with long fringes.
Pupal exuvia ( Fig. 8 View FIGURE 8 ). Pupal cases of both the sexes light brown coloured. Length of a male pupal case 2.6 mm, width 0.8 mm, in female 3.6 mm and width 1.0 mm.
Larva ( Fig. 9 View FIGURE 9 ). Mature larva 3–3.5 mm long, head dark brown, thoracal segments pale brown, more strongly sclerotized, without clearly separated darker fields. Abdomen pale yellow. Thoracic legs are well developed and blackish brown. Anal prolegs are very short.
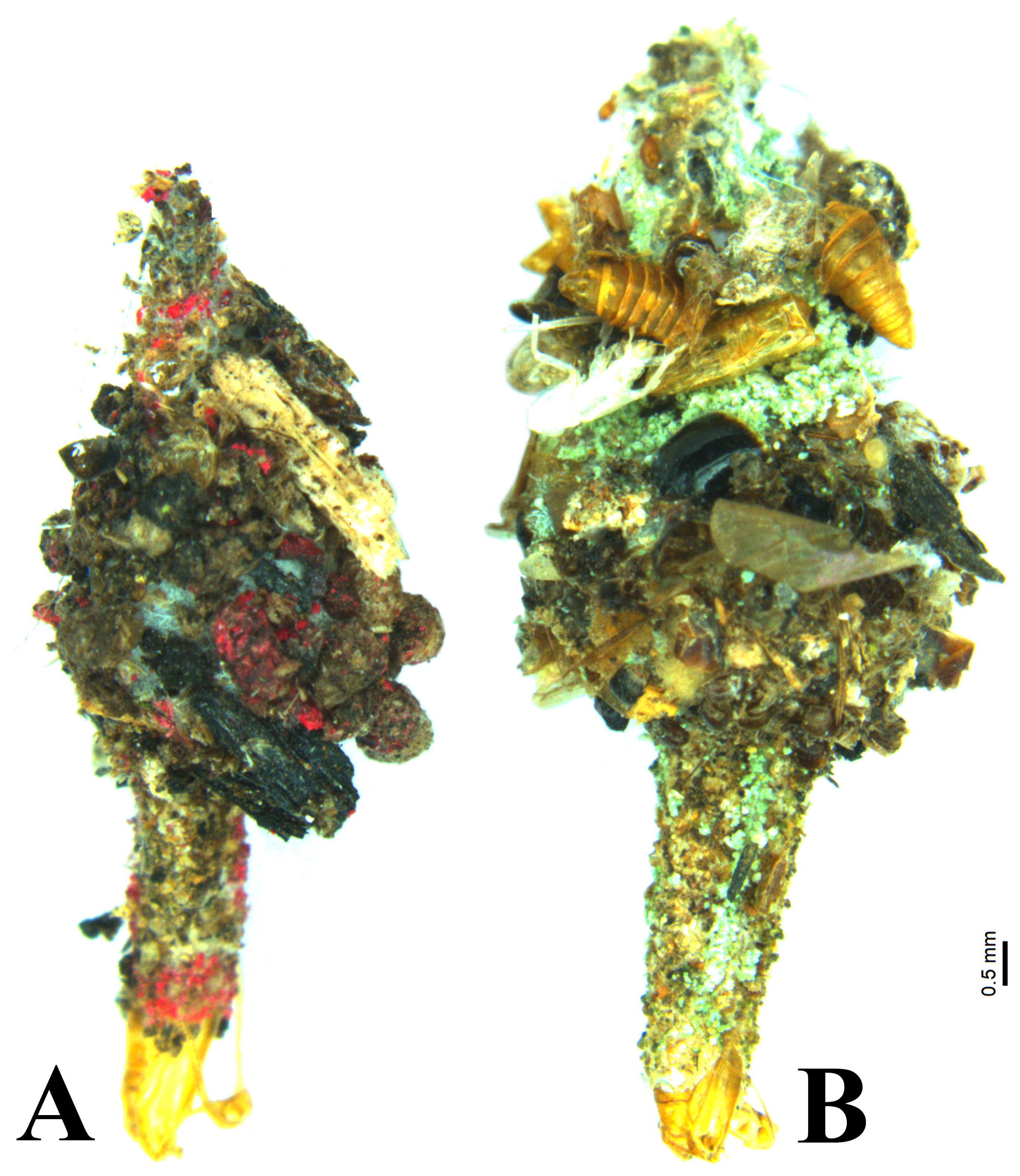
Larval case ( Fig.10a and b View FIGURE 10 ). Cases of this species are round externally with a long tube protruding at the lower end. The tube extends upward internally as long tube-like case of width 1.5 mm, it is wrapped with a sheath made up of silk, sand, soil particles, snail shells, algae, and insect debris. All the cases were found attached to the underside of the rocks with a small thread ( Fig. 11 View FIGURE 11 ). Male larval cases are smaller than females and it have 8.5–11 mm total length and 3.5–5 mm width at middle, female larval cases are 11.5–13 mm long and 4.5–6 mm wide.
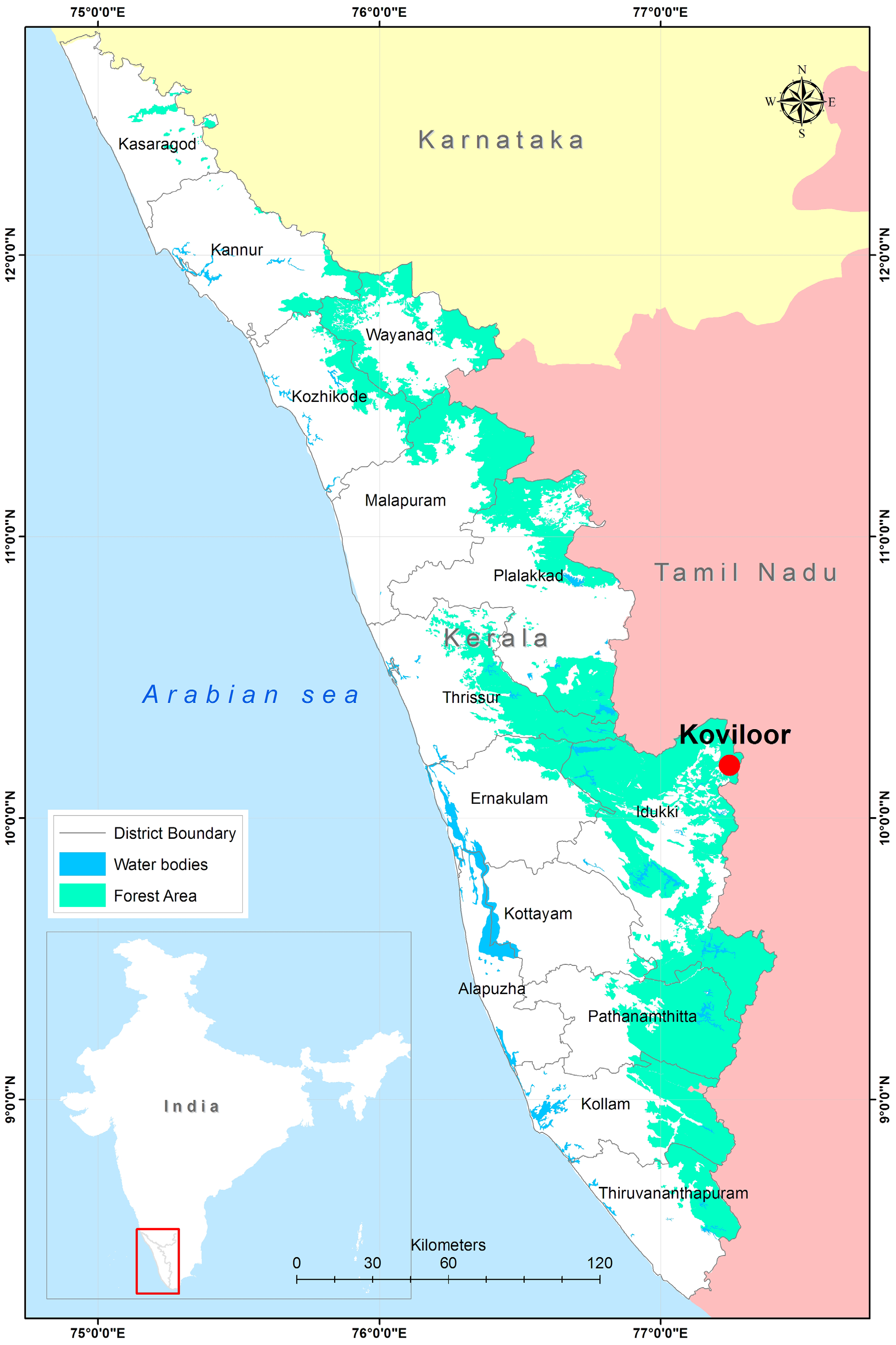
Location. The pupal cases were collected from Koviloor ( 10.18211ºN and 77.25923º E), Vattavada GoogleMaps , Idukki district GoogleMaps , Kerala. Koviloor GoogleMaps is a mixed culture village, situated in Vattavada Panchayat. It is 43.5 kms from Munnar GoogleMaps on the road to Kodaikanal GoogleMaps ( Fig. 12 View FIGURE 12 ). Altitude at the place of collection was 1700m ( 5577.42 feet) ASL. Koviloor is surrounded by different hill peaks such as Pattitaichchi mala (2497 mASL), Kaladankanava mettu (2457 mASL), Pullardi shola reserved forest, Jhanda mala (2522 mASL) and Vattavada (2153 mASL).The soil type of the region is described as ‘ Clayey mixed ustic pale humults and rock land (very deep, well drained, clayey soil on moderately steeply sloping high hills with thin vegetation, with moderate erosion, associated with rock outcrops) in Soil Maps of India (1996). In Vattavada the winter temperature ranges from 5 °C to 20 °C and in summer 12 °C to 18 °C, in monsoon season, maximum daily temperature is very low and highest temperature recorded is 19 ° C. Average rainfall according to Centre for Earth Studies Resource Atlas of Kerala (1984) is North east Monsoon – 50–60 cm; South west 150–200 cm and other rains 30–40 cm.
Biology. The larval cases were collected from underside of rocks. Active larvae were observed scrapping the surface of the rocks and it is assumed that the larvae fed on the algae and mosses on the rocks following previous reports (Saigusa and Sugimoto 2013; Roh and Byun 2016, Roh et al. 2018). Bits of algae and mosses were retrieved from the larval cases. Adult emergence was noted at night. Male moths are active flyers after emergence, they are active during day and night, whereas females moved less and stay on the case and its vicinity after emergence. It is assumed that the larger size and weight of their abdomen is a restraint to flight. Distinguishing the moths and cases from the surroundings are difficult due to it colour and positioning of cases, a characteristic suitable to evade from predators. The duration of pupation was 15–18 days. Life span of females was one week, whereas male adults lived up to 3–4 days.
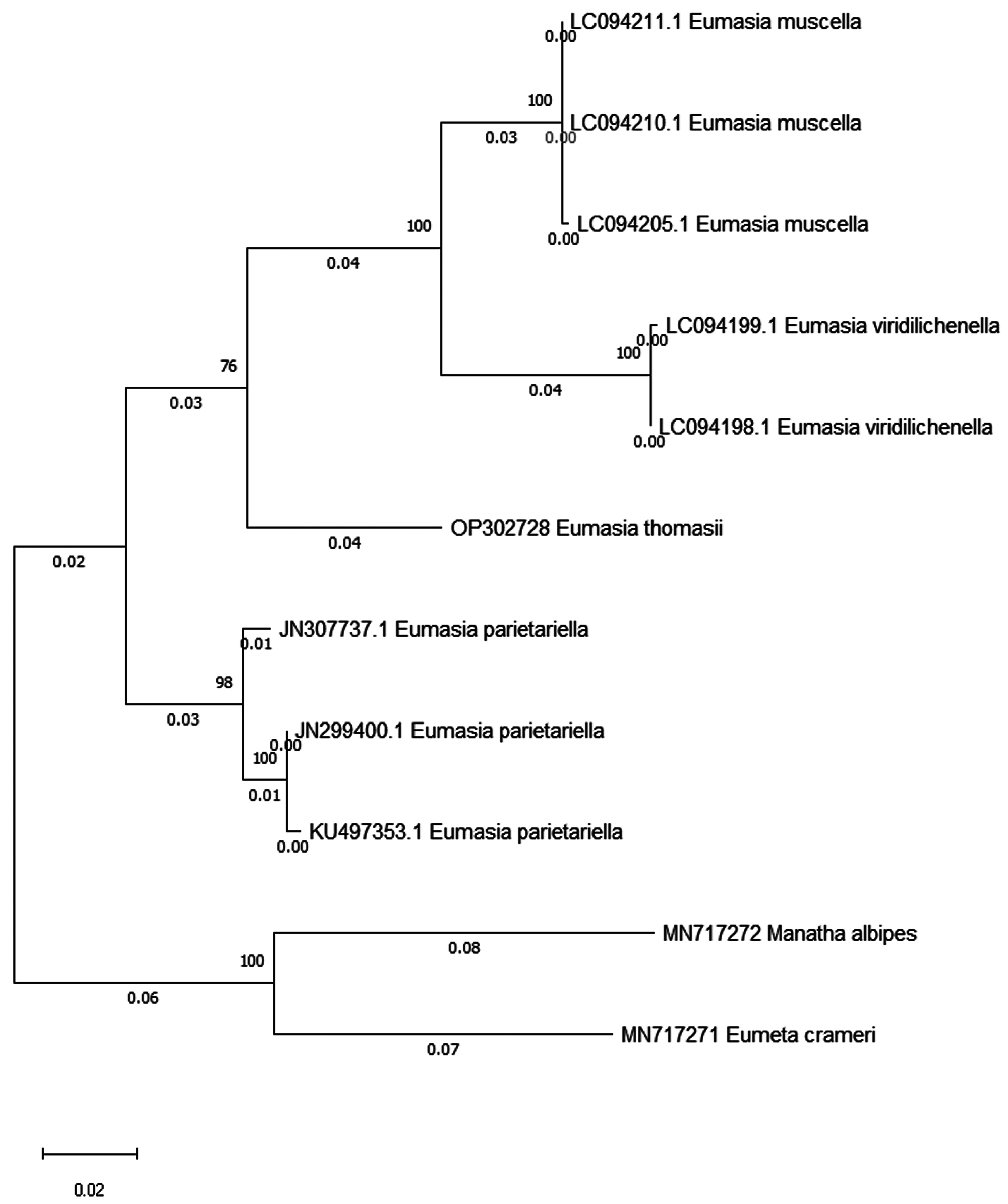
DNA barcode and phylogeny. COI sequencing of the species was done and the sequence has 687 base pairs. The sequence was uploaded in the NCBI GenBank with the accession number OP302728 View Materials . The BLAST search results of the sequence show 91.03% sequence similarity with Eumasia parietariella ( KX045802 View Materials ) from Germany and 90.83% with Eumasia muscella ( LC094210 View Materials ) from Japan .
The evolutionary relationship is depicted in the tree with the highest log likelihood (-2328.62) ( Fig.13 View FIGURE 13 ). A hypothetical taxonomic unit of Eumasia ancestor split into two branches. E. thomasii sp. nov. shows affinity to the branch having E. muscella and E. viridilichenella but seems to have evolved earlier into an independent taxon. E. parietariella belongs to the other branch. Hättenschwiler (1998) separates Eumasia into an eastern and a western group based on morphological characteristics (e.g., epiphysis on the front leg tibia, length of saccus in relation to total length of male genitalia, lobes at tegumen on male genitalia). The eastern species show an epiphysis that is absent in the western species. According to this, E. thomasii sp. nov. would belong more to the western group (with E. communita and E. parietariella ), in contrast to E. muscella and E. viridilichenella . The pronounced saccus and lobular processes of the tegumen, on the other hand, correspond to the eastern group. It seems that this division into regional groups is not consistent.
Diagnosis. Eumasia thomasii sp. nov. differs from E. muscella ( Saigusa & Sugimoto, 2005, Roh et al. 2018) from Japan and South Korea by forewing venation -, the veins R4+5 and M1, M2+3 and CuA1 are stalked in E. muscella , whereas no veins are stalked in E. thomasii sp. nov.
With regards to the difference between E. ziegleri and other Eumasia, Hättenschwiler (1998) writes: “ The species E. parvulella ( Walker, 1863) and E. arenatella ( Walker, 1864) described from Southeast Asia are larger; the latter has 8 (or 6 veins from the dc), E. testata ( Meyrick, 1911) is also larger [as E. ziegleri ], has whitish hind wings and flies March to April; E. crypsiphila Meyrick, 1919 from Sri Lanka has a similar habit but flies as early as January and E. exoria Meyrick, 1919 from Pusa, Bengal in northern India has white-grey hind wings and is larger. ” For all these species, the original descriptions say little, they are often based on individual specimens and do not allow statements about the variation.
Only two species are reported from India: E. exoria and E. arenatella . For E. arenatella , the first record from India at Kurseong, E. Himalayas, 5000 ft., in the month of September was added by Meyrick (1910) with no further information on specimen numbers or appearance: The original description by Walker gives a wingspan of 12.6 mm for the female of E. arenatella . It is thus significantly larger than E. thomasii sp. nov. E. arenatella has only six veins from the forewing dc ( Hättenschwiler 1998) but in E. thomasii sp. nov. there are seven. The flight time of E. arenatella is reported as in March, April, and September. Flight was observed in E. thomasii during January and July. E. exoria , known from the holotype male is more similar to E. thomassi . The wingspan is slightly larger than that of E. thomasii sp. nov. ( 9 mm vs. 8.0– 8.8mm). E. exoria flies later (September). The species is clearly paler. Head and thorax are ochreous-whitish ( E. thomasii sp. nov.: head golden-brown, thorax brown). The forewings of E. exoria are clearly paler and the markings stand out prominently: “ markings dark fuscous; five roundish spots on costa, two in the disc at middle and ¾ (representing stigmata), and several small elongate marks on posterior part of dorsum and termen. Cilia whitish, basal half barred dark fuscous irroration.” ( Meyrick 1919). From the middle of the posterior edge, a line sharply defined on the inside runs parallel to the outer edge towards the apex. The bright field between dc and apex of the forewings is striking (in E. thomasii sp. nov. with dark brown scales, hardly lightened).At 1/3 of the forewing, E. thomasii sp. nov. a slanting, dark band of scales from anterior to posterior edge. In E. exoria , the basal half of the wing is clearly brighter and almost without markings. An overview of the Asian species ( Table 3 View TABLE 3 ) summarizes the differences of these species from E. thomasii sp. nov. based on available information.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Eumasiinae |
|
Genus |