
Anhanguera adductor subsp. mandibulae
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlaa163 |
|
persistent identifier |
https://treatment.plazi.org/id/AB191354-7A33-D467-FF47-41C085F5F9E8 |
|
treatment provided by |
Plazi |
|
scientific name |
Anhanguera adductor subsp. mandibulae |
| status |
|
M. adductor mandibulae externus superficialis (mAMES)
Origin: mAMES is the lateralmost of the temporal muscles, which originates from the temporal bar – more precisely from the lateral surface of the squamosal and ventromedial surface of squamosal and postorbital in lepidosaurs, from the squamosal and quadrate otic process in modern birds, and from the anterolateral surface of the quadrate and quadratojugal in crocodylians ( Holliday & Witmer, 2007). The configuration in crocodylians is most likely shifted from the squamosal as a derived feature, given its origin on the squamosal in both lepidosaurs and modern birds, while the absence of this muscle on the surface of the postorbital in modern birds is likely due to the loss of the temporal bar in these animals. The conservative morphology of the temporal bar in pterosaurs, comprising both the postorbital and the squamosal, most likely indicates that these two bones served as origin areas for mAMES. Unfortunately, the origin of this muscle is usually fleshy and rarely leaves osteological correlates.
Insertion: mAMES runs lateral to the skull and inserts on the lateral surface of the mandible in modern birds, on the dorsolateral surface of the surangular in lepidosaurs, on the dorsal surface of the surangular in crocodylians ( Holliday & Witmer, 2007) and most likely on the dorsolateral surface of the angular in non-avian dinosaurs ( Holliday, 2009). Insertion sites for this muscle generally do not leave clear osteological correlates, only rarely leaving smooth shallow fossae ( Holliday & Witmer, 2007). Considering the phylogenetic bracketing, this muscle would most likely insert dorsolaterally on the surangular of pterosaurs.
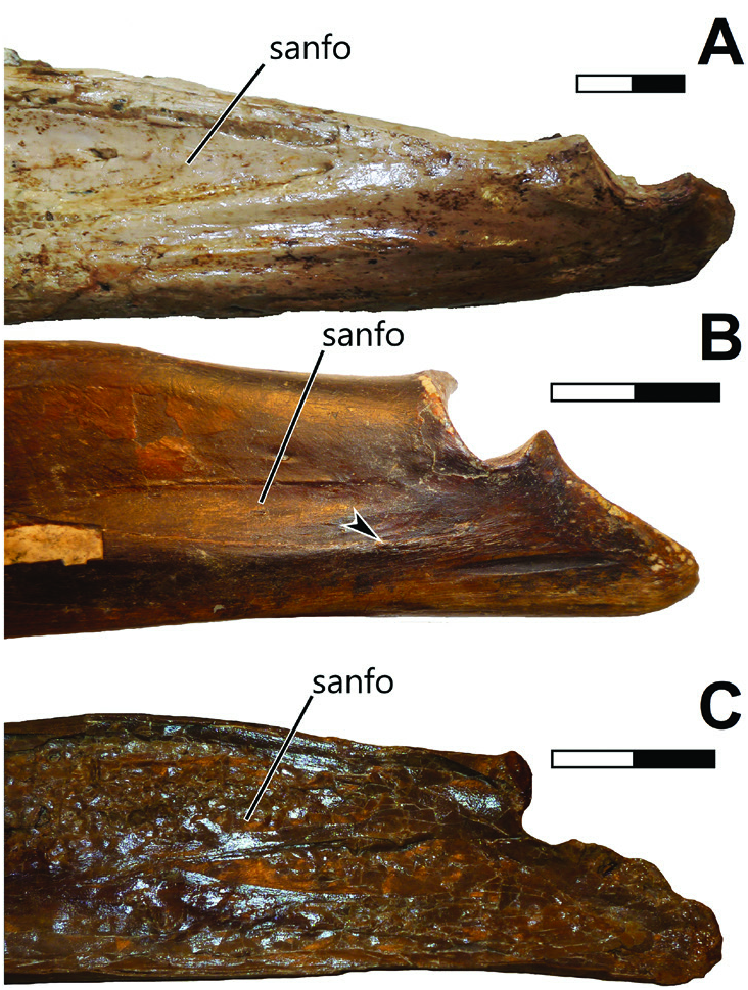
Extensive scarring can be seen in the dorsal surface of the surangular of An. araripensis and pteranodontids ( Fig. 16 View Figure 16 ), which seems to correlate at least partially to mAMES. The lateral surface of the bone is gently excavated by a depression, which extends further ventrally to part of the angular. This depression bears some faint muscle scars. A similar depression can be seen in the holotypes of Th. sethi and Ludodactylus sibbicki ( Fig. 17 View Figure 17 ), although is the latter specimen the surface is damaged and no scars can be seen except on the dorsal region of the surangular. This surangular/ angular depression is situated dorsal to the probable insertion site of mPTv, which was previously discussed. As mentioned, in An. araripensis , a strong ridge seems to separate the insertion areas of these two muscles.
ESTIMATED BITE FORCES AND BFQ
Reconstructed CSAs ( Fig. 2 View Figure 2 ), inlevers, outlevers, mechanical advantages and estimated bite forces are given in Tables 3–4. The values of stylopodial diameters are given in Table 5, and estimated body masses and BFQs are given in Table 6.
Considering the mean values of bite force (tip and 70% jaw-length), the lowest bite forces were found in the two smallest specimens: 8.67– 12.40 N (tip and 70% jaw-length, respectively) for Ta. wellnhoferi MN 6595-V ( 2.23 kg in body mass) and 5.0– 7.9 N for Nyctosaurus gracilis FMNH P 25026 ( 3.2 kg). The same values for D. weii IVPP V 4065 were 61 – 86 N, despite being only slightly larger than the former two ( 4.18 kg). The mean mid-jaw length bite force calculated for D. weii , due to its unique dentition, was 185 N. Pteranodon longiceps YPM 1177 ( 15.12 kg) and An. blittersdorffi MN 4805-V ( 10.45 kg) bear similar mean tip and 70% jaw-length bite forces (~23–~33 N), although the estimated bite forces of the similar-sized C. ybaka MN 4726-V ( 13.61 kg) were over twice as high (~52–74 N), close to the estimated bite forces of Tr. mesembrinus BSPG 1987 46 (58–82 N), which is larger than the former three (~ 23 kg). The two largest analysed specimens, Th. sethi DGM 1476 -R and Tupuxuara leonardii IMNH 1052 (~ 30 kg), exhibit disparate estimated bite forces: 157–224 N and 29–41 N, respectively.
The largest BFQs were estimated for D. weii (26.88) and Th. sethi (20.61), followed by Ta. wellnhoferi (13.55), C. ybaka (9.81) and Tr. mesembrinus (8.68). Estimated values were lower for the other taxa: Tupuxuara leonardii (3.94), N. gracilis (3.18), Pteranodon longiceps (4.04) and An. blittersdorffi (5.14).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.