
Ectenoglossa, Sinclair, 1945
|
publication ID |
https://doi.org/10.11646/zootaxa.3076.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/A87D878B-FFE2-FFED-0BA8-FABFF98AFBE4 |
|
treatment provided by |
Felipe |
|
scientific name |
Ectenoglossa |
| status |
|
Ectenoglossa ? oviforma sp. nov.
Pl. 1, Figs. 13–18; Pl. 2, Figs. 1 View FIGURE 1 –5, Table 2
1975 Ectenoglossa ? sp.―Krause & Rowell, pp. 27–28, Pl. 3, Figs. 20–23.
Derivation of name. Refers to the egg-shaped outline.
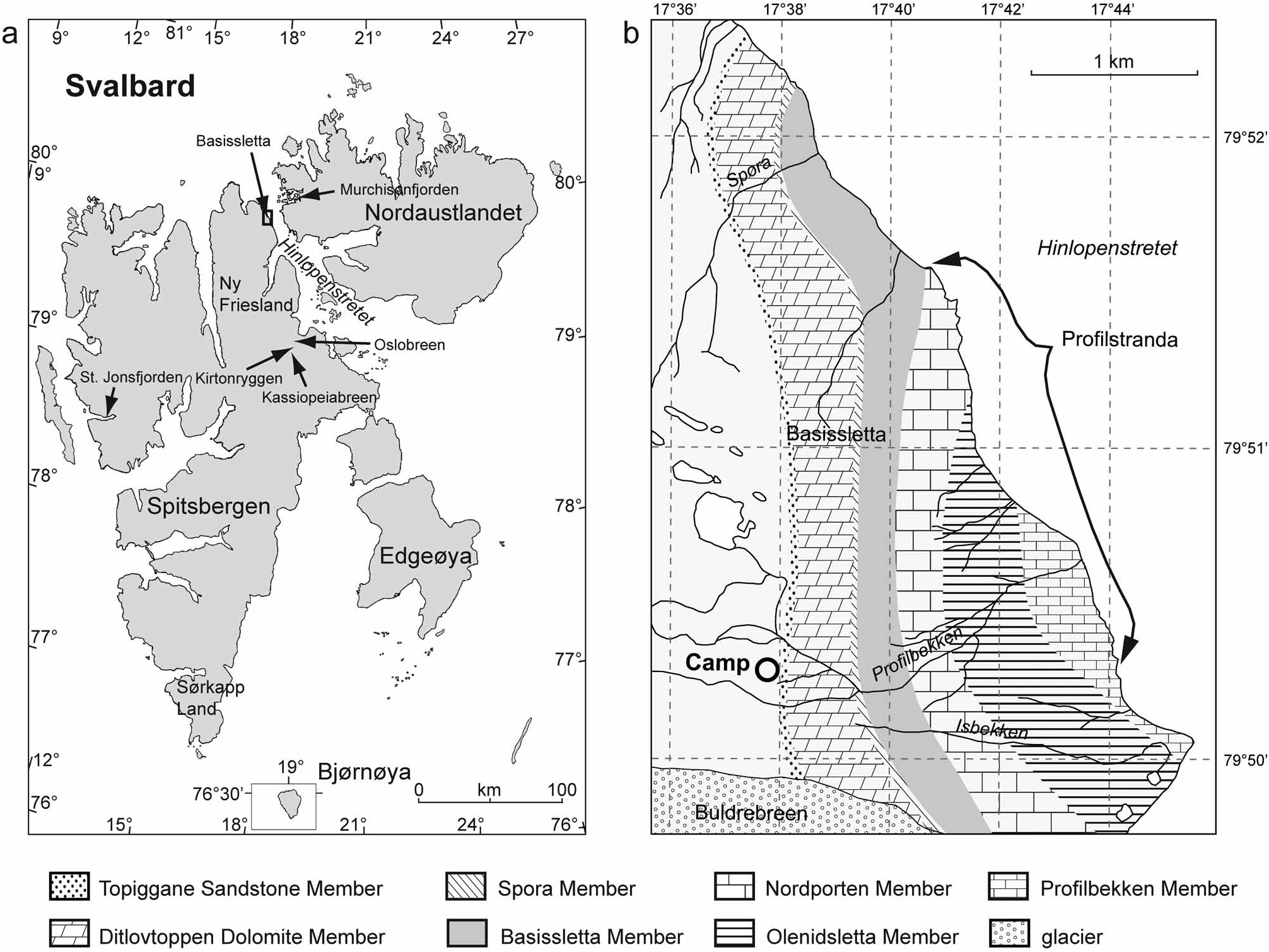
Holotype. Pl. 2, Fig. 2 View FIGURE 2 ; F3028/3678, ventral valve; 85 m above base of Olenidsletta Member, Valhallfonna Formation, sample F3028/F3678; Profilstranda at Basissletta, Ny Friesland, Spitsbergen.
Material. 405 specimens from samples with A84137 View Materials , A84162 View Materials , A84165 View Materials , A84306 View Materials , A84363 View Materials , A84364 View Materials and A84382 View Materials and from samples F3028, F3173, F3503, F3533, F3546, F3560, F3574, F3596, F3664, F3667, F3678, F3680, F3777, F3817, F3828, F3839, F3840, F4721, F5043, F5155, F5188, F5758, JH-6, JH-12, JH-14–18, JH-21, JH-23, JH-26, JH-27, JH-30, JH-32, JH-33, JH-35, JH-37, JH-39, JH-40, JH-42–44, JH-46–49, JH-56, JH-57, JH- 75, JH-95, JH-96, JH-98, JH-99, JH-104, JH-108, JH-111, JH-113, JH-122, JH-129, JH-130, JH-133, JH-136, JH- 139, JH-140, JH-155, JH-164, JH-168, JH-178, JH-182, JH-189 and JH-191–195. The paratypes are F3840, TSGF16933–16935 , TSGF17003–17005 , TSGF17080 and TSGF17081 .
Diagnosis. Elongate, egg-shaped or rounded spatulate Ectenoglossa with L/W ratio 1.3–2.1; profile generally slightly biconvex; thin dorsal median ridge reaching slightly anterior to mid-valve; short ridge connecting pedicle groove to valve floor.
Description. Shell large, spatulate to elongate egg-shaped, slightly ventribiconvex in longitudinal profile and slightly to moderately (rarely strongly) convex in cross-section. Anterior valve margin generally flattened in larger specimens but evenly rounded in small specimens. Shell generally without shoulders and widest at 41–69% of valve length. Valves about 10–13% as deep as they are long and deepest at 30–33% of valve length. L/W ratio 1.12–1.99 ( one specimen exceeding 2.11), increasing with size. Largest specimen 38.2 mm long and 24.6 mm wide. Shell thin-walled. Growth lines dominate ornamentation, often developing into thin, low rugae or filae. Rugae 5–18 per mm. Fine radial striation on some specimens. Micro-ornamentation absent.
Dorsal pseudointerarea anacline, 0.5–2% as long as valve and well defined but not raised substantially above valve floor. Propareas divided by wide, crescent-shaped, shallow to moderately deep median groove. Proximal part of visceral area with scattered large, circular pits, even in a specimen no wider than 3.6 mm. Pits decreasing in size and density distally in visceral area. Diameter of larger pits about 0.4 mm. Distribution of pits outside visceral area variable, but large, rather dense pitting occurring in a band along lateral valve margin and normally also bordering anterior margin of visceral area. Well-preserved specimens show dense, very fine pitting between larger pits. Valve floor generally striate with obscure visceral area. Dorsal median ridge thin and weak when developed, reaching 52– 61% of valve length. Muscle scars generally obscure. Central muscle scars subcircular or transversely oval, about 5–7% as long as valve and located at 40–50% of valve length. Anterior lateral muscle scars located at 55–60% of valve length. Gastroparietal bands strongly diverging and impressed about one muscle length posterolaterally to central muscle scars. Baculate mantle canal system often well impressed with peripheral vascula lateralia and strongly branching vascula media. Two branches of vascula lateralia converging from about 40–50% of valve length to well anterior of visceral area, then diverging again. Vascula media subparallel along median ridge but diverging from anterior point of ridge and branching at about 80% of valve length.
Ventral pseudointerarea apsacline to orthocline, about 43–55% as wide as valve, often declining toward valve floor but separated therefrom. Pseudointerarea 1–6% as long as valve. Flexure lines rather weak, nearly straight or bending anteriorly, defining small triangular inner parts of propareas. Pedicle groove moderately deep and sharply defined, widening from 7–8% of valve width to 9–10% of valve width. Pedicle groove connected to valve floor by short ridge extending anteriorly from groove. Pitting and striation of valve floor like that in dorsal valve. Visceral area generally with weakly impressed muscle scars. Pedicle nerve impressions reaching 16–61% of valve length, diverging at about 5˚, but sometimes diverging more in proximal part. Three weak ridges reaching 12% of valve length. Middle lateral muscle scars oblique, oval, reaching about 45–50% of valve length. Vascula lateralia of baculate mantle canal system running about 20% of valve width from lateral valve margin forward to about 35–50% of valve length, then turning in an anteromedian direction. Branches of vascula lateralia becoming subparallel when they reach about 40% of valve width from valve edge.
Remarks. Ectenoglossa is poorly understood because the type species, E. lesueuri Rouault , is represented by poorly preserved specimens. For the same reason, the specimens from Spitsbergen are questionably assigned to the genus. Although the genus has been provisionally assigned to the Glossellinae ( Holmer & Popov, 2000) it is here considered to belong within the Obolinae in view of the presence of a dorsal pseudointerarea in the new species. One of the authors (Holmer) has examined the types of the type species and found that the absence of a dorsal pseudointerarea may be preservational.
Ectenoglossa ? oviforma sp. nov. shows some morphological changes up through the succession; stratigraphically younger specimens are generally larger, more elongate and more transversely convex than older ones, giving the impression that there are two separate species. However, the large amount of material available does not support this impression.
The Ectenoglossa ? sp. described and figured by Krause & Rowell from the basal part of the Middle Ordovician (Dapingian) Antelope Valley Limestone in Nevada appears to have the same diagnostic features as specimens from the upper part of the Valhallfonna Formation. The specimens from Nevada are therefore included in E.? oviforma. The species E. angusta Yadrenkina, 1977 from the Darriwilian of Siberia has strong, radiating ridges on the distal part of the exterior and shows a rounded umbo. Another species from the Darriwilian of Siberia, E. derupta Yadrenkina, 1977 , has less developed coarse pitting of the valve floor and a pentagonal outline. The Middle Ordovician Russian species E.? lata ( Pander, 1830) is angular and wide. The type species, E. lesueuri ( Rouault, 1850) is distinctly pentagonal, with rather long and strongly developed subparallel ridges on the posterior part of the ventral valve and a very poorly developed dorsal median ridge. The Darriwilian E.? lyelli ( Billings, 1859a) from Canada differs in having subparallel lateral margins of the shell, moderately convex valves and a rather high L/W ratio for its size. E. magna Gorjansky, 1972 is large, lacks dorsal pseudointerarea and has orthocline ventral pseudointerarea. The species E. minor Zhan & Cocks, 1998 from the Ashgill of China has an L/W ratio of 2.6–2.8 and is small. The Upper Ordovician E. missouriensis Rowley, 1908 from Missouri is large and has an obtuse umbo, parallel sides, and a shallow and narrow sulcus. E. nympha (Billings, 1865) from the Middle Ordovician of Newfoundland has a very high L/W ratio for its size and subparallel sides. E. nymphoidea Cooper, 1956 from the Sandbian of the USA is distinguished by its large size, moderate convexity and poorly defined mantle canal system. E. philomela (Billings, 1862) from the Katian in eastern Canada is easily distinguished by a high L/W ratio for its size, a sulcus on both valves, and strong radiating striae on the valve floor. E.? rubra Cooper, 1956 from the Sandbian of Tennessee, USA, is distinguished by its strongly diverging posterior-valve margins, moderate convexity and subparallel lateral margins. E.? sculpta Cooper, 1956 from the Darriwilian of Oklahoma, USA, is moderately convex with a narrower, more rounded anterior margin and a more distinct median ridge. The Upper Ordovician E. sorbulakensis Popov, 1980 from Kazakhstan is markedly more elongate compared to its size and has a deep pedicle groove, welldefined pedicle nerve impressions, and no dorsal pseudointerarea. Cooper (1956) also described Ectenoglossa sp. 1 from the Middle Ordovician of Newfoundland, which differs in the obtuse angle of the umbo and may differ in the evenly rounded anterior margin.
Occurrence. 75–101, 108–110 and 135–145 m above base of Olenidsletta Member and 17–21, 25–29, 40, 45– 52, 61, 67–72, 81 and 98 m above base of Profilbekken Member, Valhallfonna Formation, Basissletta in northeastern Ny Friesland, Spitsbergen.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.