
Handleyomys intectus ( Thomas, 1921 )
|
publication ID |
https://doi.org/10.1093/mspecies/sez007 |
|
publication LSID |
lsid:zoobank.org:pub:72734727-F885-40D0-83FD-A935CEC9BFAD |
|
persistent identifier |
https://treatment.plazi.org/id/A763247E-FF87-DD19-BDC9-1C4D338EFB28 |
|
treatment provided by |
Felipe |
|
scientific name |
Handleyomys intectus ( Thomas, 1921 ) |
| status |
|
Handleyomys intectus ( Thomas, 1921) View in CoL
Colombian Rice Rat
Oryzomys intectus Thomas, 1921:356 View in CoL . Type locality “ Santa Elena, ” [ Colombia], extended to “ Colombia, Antioquia Dept., 8 km E Medellín, Santa Elena, 9000 ft ( 2743 m),” by Musser and Carleton (2005:1,118).
[ Oryzomys ( Oryzomys) ] intectus: Tate, 1932:16 View in CoL . Name combination.
Nectomys View in CoL [ intectus View in CoL ]: Ellerman, 1941:351. Name combination.
Handleyomys intectus: Voss, Gómez-Laverde and Pacheco, 2002:5 View in CoL . First use of current name combination.
CONTEXT AND CONTENT. Context as for genus, no subspecies are recognized (Gómez-Laverde et al. 2015).
NOMENCLATURAL NOTES. Thomas’s (1921) original description of the species was based on the holotype (BMNH [British Museum of Natural History] 21.7.1.17, preserved as a skin and skull) and two paratypes, all of which were collected at Santa Elena, near Medellín, Colombia, by Nicéforo María in December 1919 and January 1920 ( Voss et al. 2002). The generic name was given in honor of Charles O. Handley, Jr. for his contributions to Neotropical mammalogy ( Voss et al. 2002).
The genus Handleyomys was separated from Oryzomys based on several characteristics: Oryzomys species have abruptly countershaded head and bodies, and bicolored tails, whereas Handleyomys lack countershading and have dark, unicolored tails. The plantar surface of the hind foot is covered with squamae and the lateral tarsal or hypothenar pad is absent in Oryzomys ( Voss et al. 2002) . Handleyomys was separated from the ex “ Oryzomys alfaroi ” group taxa based on the number of mammae (6 versus 8 in the alfaroi group— Weksler 2015). The alfaroi group was tentatively included as “ Handleyomys ” because no new genus to encompass these morphologically and genetic distinctive taxa has been described ( Weksler 2015). Based on morphological data, H. intectus has been associated with the Andean Oreoryzomys balneator , Melanomys , or Nectomys (Musser and Carleton 2005) , but these taxa are not closely related to Handleyomys . Several morphological and molecular phylogenetic analyses recovered Handleyomys as a wellsupported clade (e.g., Weksler 2003, 2006; Almendra 2015). English common names are white-footed Handley’s mouse, Colombian Central Andes cloud forest mouse, or Colombian rice rat (Musser and Carleton 2005; Gómez-Laverde and Delgado 2008; Gómez-Laverde et al. 2015); a local name used is rata arrocera antioqueña (Rodriguez-Mahecha et al. 1995).
DIAGNOSIS
The hindfeet of Handleyomys intectus are usually covered dorsally by pure-white (or indistinctly pigmented) hairs, whereas the hindfeet of the dusky-footed Handley’s mouse H. fuscatus are covered dorsally with dark-banded hairs. The nasals usually extend posteriorly well beyond the premaxillary-maxillary suture in H. intectus , whereas in H. fuscatus the nasals are usually truncated posteriorly at or near the premaxillary-maxillary suture. The incisive foramina usually have smoothly tapering (never abruptly constricted) lateral margins in H. intectus , whereas in H. fuscatus the incisive foramina have lateral margins that are usually constricted abruptly anteriorly. Transversally, the interparietal is narrow in H. intectus , whereas in H. fuscatus it is wider ( Voss et al. 2002).
GENERAL CHARACTERS
Handleyomys intectus is a medium-sized terrestrial rat ( Fig. 1 View Fig ). Adult pelage is fine and soft, dull brownish-gray dorsally, usually darker middorsally than on the flanks. The ventral pelage is dark gray frosted with paler gray or buff. The tail is unicolored (dark) and usually as long as the combined length of head and body. The manus are sparsely covered dorsally with short pale (whitish or silvery) hairs; ventral surface naked and unpigmented, with five separate plantar tubercles (two carpal and three interdigital pads). Pes are long and narrow, with digits I and V much shorter than middle three, with six plantar pads (two metatarsal and four interdigital pads); conspicuous ungual tufts of long silvery hairs rooted at bases of claws on second through fifth digits, but pedal dorsum otherwise only sparsely covered with short pale or dark-banded hairs. Six mammae are present in inguinal, abdominal, and postaxial pairs ( Voss et al. 2002). The presence of six mammae is an unusual feature within oryzomyines, which generally have eight mammae ( Weksler 2006).
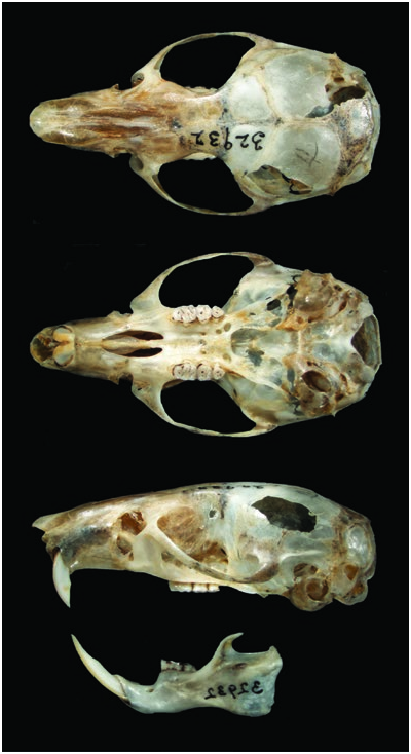
The skull of H. intectus ( Fig. 2 View Fig ) presents a long and tapering rostrum flanked by shallow but distinct zygomatic notches; interorbital region hourglass-shaped; braincase is rounded and moderately inflated, lacking prominent temporal crests, ridges, or beads. Zygomatic plate moderately broad with a vertical anterior edge. Premaxillae short, not forming a rostral tube. Palatal bridge wide and long, with large posterolateral pits. Mesopterygoid fossa not fenestrated. Postglenoid foramen separated from the large subsquamosal fenestra by a slender hamular process of the squamosal. Tegmen tympani without or with only shallow squamosal overlap. Pars flaccida of the tympanic membrane large. Orbicular apophysis of the malleus well-developed. Bullae small ( Voss et al. 2002; Gómez-Laverde et al. 2015).
Means, ranges, and n of external and skull measurements (mm) and masses (g) of adult H. intectus from the departments of Antioquia, Quindío, and Risaralda in Colombia were ( Voss et al. 2002): head and body length, 101 (85–114; 47); length of tail, 95 (75–106; 47); length of foot, 24 (21–26; 50); length of ear, 14 (10–17; 43); condylo-incisive length, 23.8 (21.0–25.5; 44); length of diastema, 7.2 (5.9–7.9; 50); occlusal length of the maxillary molar row, 4.1 (3.8–4.4; 51); breadth of the first maxillary molar, 1.3 (1.2–1.4; 51); length of one incisive foramen, 4.5 (3.8–5.0; 50); breadth across both incisive foramina, 2.0 (1.7– 2.2; 50); breadth of the palatal bridge, 2.7 (2.2–3.1; 50); breadth of the zygomatic plate, 2.3 (1.8–2.6; 50); least interorbital breadth, 4.9 (4.6–5.3; 51); zygomatic breadth, 14.0 (12.8–14.9; 42); nasal length, 11.2 (9.5–12.5; 45); interparietal breadth, 7.7 (6.2–8.8; 46); and body mass, 26 (17–33; 25).
The maxillary molars incipiently lophodont. The maxillary molar rows are parallel. The principal molar cusps are bunodont when unworn; the labial and lingual re-entrant folds are interpenetrating. M1 anteromedian flexus absent. M3 subtriangular in occlusal view, smaller than M1 and M2. Incisors ungrooved and opisthodont. Lower incisors alveoli lack capsular process on lateral mandibular surface ( Voss et al. 2002; GómezLaverde et al. 2015).
Available samples of H. intectus are remarkably similar in size throughout the known range of the species. The only noteworthy exception to this tendency is the interparietals of specimens from Quindío and eastern Risaralda, which are markedly narrower, on average, than those of specimens collected in Antioquia ( Voss et al. 2002).
Means (ranges; n) of external measurements (mm) and masses (g) of adult females from Antioquia, Colombia were (data collected by Carlos A. Delgado-V. and LMV): total length, 178.0 (164–186; 6); length of tail, 84.8 (77–92; 6); length of foot, 22.6 (21.2–23.3; 6); length of ear, 13.2 (11.6–13.8; 6); and body mass, 24.0 (19–34; 6). Means (ranges; n) of external measurements (mm) and masses (g) of adult males from Antioquia, Colombia were (data collected by Carlos A. Delgado-V. and LMV): total length, 185.6 (163–204; 22); length of tail, 90.2 (75.0–103.9; 22); length of foot, 23.3 (21.3–25.7; 22); length of ear, 14.0 (11.5–16.0; 22); and body mass, 24.1 (17–30; 21).
DISTRIBUTION
Handleyomys intectus is an endemic Andean species ( Fig. 3), distributed in the Cordillera Central of Colombia over an elevational range from 1,500 to 2,800 m in the departments of Antioquia, Quindío, Risaralda, and Valle del Cauca ( Voss et al. 2002; Rojas- Díaz et al. 2012). The locality El Roble was reported as occurring in the department of Cauca ( Weksler 2006) when, in fact, it is located in the department of Quindío (see Voss et al. 2002). No fossils are known.
FORM AND FUNCTION
Mystacial, superciliary, genal, submental, interramal, and carpal vibrissae are present. Pinnae not large but clearly visible above fur of head, sparsely covered with short dark hairs. Tail about as long as combined length of head and body, apparently naked (a sparse caudal pelage is only visible under magnification), and unicolored (dark above and below). The carotid arterial morphology is primitive, with orbitofacial circulation supplied by separate supraorbital and infraorbital branches of large stapedial artery (pattern 1 of Voss 1988); course of supraorbital stapedial ramus marked by prominent squamosal-alisphenoid groove and sphenofrontal foramen ( Voss et al. 2002). The stomach is unilocular and hemiglandular, without any extension of glandular epithelium into corpus; bordering fold crosses lesser curvature slightly to right of incisura angularis; bordering fold crosses greater curvature opposite incisura angularis. Gall bladder is absent ( Voss et al. 2002). The morphology of male reproductive structures including the glans penis is unknown (GómezLaverde et al. 2015).
Dental formula is: i 1/1, c 0/0, p 0/0, m 3/3, total 16 ( Voss et al. 2002). Voss et al. (2002) and Gómez-Laverde et al. (2015) describe the dentition in great detail.Anterocone of M1 not divided into labial and lingual conules. Anteroflexus very deep, extending lingually beyond the dental midline on M1 and M2. Anterolophs and mesolophs large, fused with corresponding styles on the labial margins of M1 and M2. Posterolophs distinct on M1 and M2, persisting with moderate to heavy wear. M1 has one accessory labial root (four roots total), but M2 and M3 are each three-rooted. Anteroconid of m1 undivided by a median flexid and fused with the protolophid or the anterolophid (sometimes both) to enclose a persistent internal fold of uncertain homology (might be anteroflexid, protoflexid, or both). Anterolophid absent on m2 and m3.Anterolabial cingulum absent or indistinct on m2, and absent on m3. Mesolophids and posterolophids large and well developed. Ectolophids absent. The m1 with an accessory labial root and occasionally also an accessory lingual root ( 3–4 in total); m2 and m3 each have three roots, two small and anterior and one large and posterior (Gómez-Laverde et al. 2015).
Vertebral formula is 7C, 19 TL, 4 S, 28–30 Ca, total 58–60. The caudal vertebrae lack hemal arches. Twenty-four ribs are present. The tuberculum of first rib articulates with transverse processes of seventh cervical and first thoracic vertebrae; second thoracic vertebra with greatly elongated neural spine; entepicondylar foramen of humerus absent ( Voss et al. 2002).
ONTOGENY AND REPRODUCTION
A pregnant female who gave birth (two offspring) in a trap was caught in the month of April during the rainy season, whereas one lactating female was caught during the month of January during the dry season. Juveniles have been found in both seasons (Gómez-Laverde et al. 2015).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Handleyomys intectus ( Thomas, 1921 )
| Vargas, Laura M, Ramírez-Chaves, Héctor E & Velazco, Paúl M 2019 |
Nectomys
| ELLERMAN, J. R. 1941: 351 |
Oryzomys ( Oryzomys )
| TATE, G. H. H. 1932: 16 |
Oryzomys intectus
| THOMAS, O. 1921: 356 |