
Calumma boettgeri
|
publication ID |
https://doi.org/10.11646/zootaxa.4048.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:2CAB0746-175E-4FE1-B0D0-23DFF395A559 |
|
DOI |
https://doi.org/10.5281/zenodo.5619475 |
|
persistent identifier |
https://treatment.plazi.org/id/A1328788-9B6D-FF9D-FF38-F9D2FA17F946 |
|
treatment provided by |
Plazi |
|
scientific name |
Calumma boettgeri |
| status |
sensu lato |
Morphology of Calumma boettgeri sensu lato
External morphology. Measurements of important morphological parameters were taken from 23 specimens ( 11 males, 12 females) from Nosy Be, six specimens (five males, one female) from Montagne d’Ambre, and another nine specimens without exact locality data ( Table 1). Because there was only one female from Montagne d’Ambre available, only males from both species were considered for comparison of body size, extremities and appendages. The specimens without localities were not included in mean value calculations but could be assigned according to their morphology to the Nosy Be morphotype (four specimens) and the Montagne d’Ambre morphotype (five specimens), respectively.
, ratio of tail to snout-vent length; LRA, length of rostral appendage from snout tip; RRS, ratio of length of rostral appendage and snout-vent length; RAPSC, number of peripheral scales on rostral appendage; NPSCM, number of peripheral scales per mm on rostral appendage; RC, rostral crest present (+) or absent (-); NSL, number of supralabials; NIL, number of infralabials; OLD, lateral diameter of the occipital lobe; OLN, depth of the dorsal notch in occipital lobe; PC, parietal crest absent (-) or number of parietal cones; DC, dorsal crest absent (-) or number of dorsal cones; AP, axillary pits present (+) or absent (-); DSC, diameter of broadest scale on upper arm; RSB, ratio of broadest scale to snout-vent length; NSC, number of big scales on upper arm from lateral view; UAD, upper arm diameter; RAS, ratio of arm diameter to snout-vent length; all measurements in mm.
……continued on the next page …….continued on the next page Continued.
collection no. species locality sex NIL OLD OLN PC DC AP DSC RSB NSC UAD RAS ……continued on the next page collection no. species locality sex NIL OLD OLN PC DC AP DSC RSB NSC UAD RAS SMF 16471 C. boettgeri Nosy Be f 10 4.2 0.0 - 0 - 0.4 0.007 9 2.2 0.044 SMF 16472 C. boettgeri Nosy Be f 13 3.5 0.1 - 11 - 0.4 0.009 10 1.8 0.039 ZSM 865/1920 C. boettgeri - f 13 2.8 0.1 - 0 - 0.3 0.006 9 1.9 0.038 ZFMK 51515 C. boettgeri - f 13 3.3 0.7 - 0 - 0.3 0.006 12 2.1 0.041 MCZ 5988 Ch. macrorhinus ‘Madagascar’ f 6.1 3.2 0.1 - 0 - 0.3 0.007 8 2.3 0.047 Individuals from Montagne d’Ambre show clear morphological differences from Nosy Be specimens ( Fig. 7 View FIGURE 7 ). The adult males examined from Montagne d’Ambre are larger than those from Nosy Be (mean values of TL 107.8 mm in Montagne d’Ambre vs. 99.3 mm in Nosy Be, Table 2 View TABLE 2 ), their rostral appendage is longer related to the snoutvent length (RRS 0.078 vs. 0.065) and the ratio of arm diameter to snout-vent length is larger (UAD/SVL 0.053 vs. 0.042).
species C. boettgeri C. boettgeri C. linotum
locality Nosy Be Nosy Be M. d'Ambre
sex m f m
Both populations show heterogeneous scalation, especially at the extremities and the rostral appendage. The enlarged rounded tubercles on the limbs are distinctly larger in animals from Montagne d’Ambre; the mean diameter of the largest tubercle of the upper arm is 0.64 mm compared to 0.36 mm ( Nosy Be). Additionally the number of the enlarged tubercles on the upper arm of males from Montagne d’Ambre is approximately twice that of males from Nosy Be, with a mean of 20.2 compared to 9.9, respectively, and the tubercles are not bordering each other on Nosy Be individuals. This character is also confirmed from a juvenile of SVL 26.9 mm (ZFMK 48227, Nosy Be) and a juvenile of SVL 36.2 mm (ZFMK 52308, Joffreville). Another morphological difference between both populations can be found in the pholidosis of the rostral appendage. Although the rostral appendage is significantly smaller in males from Nosy Be (see above), they have more peripheral scales on it, with a mean of 18.9 compared to 15.5 in Montagne d’Ambre males. In relation to the size of the appendage, this means 6.1 scales per mm compared with 4.1 scales per mm in Montagne d’Ambre. In summary, specimens from Montagne d’Ambre show a more heterogeneous scalation with broader tubercles on extremities and the rostral appendage. This is true of females as well (note that only one female with locality data was examined). Montagne d’Ambre individuals differ also in both sexes from Nosy Be specimens in the presence of a parietal crest which is best visible in the micro-CT scan (see below).
The other morphological features either were highly variable or did not differ between the populations. For example, the number of dorsal cones was 0–28 in Nosy Be, and 9–13 in Montagne d’Ambre. Likewise the edges of the occipital lobes vary between the specimens. They range between un-notched and clearly notched (up to 0.7 mm) in Nosy Be, and are either not or only slightly notched (up to 0.3 mm) in Montagne d’Ambre. All specimens have a rostral crest and none have axillary pits.
Colouration. Males also show great differences between the populations in colouration (see Fig. 2 View FIGURE 2 A, B). Males from Montagne d’Ambre were more colourful, with a true blue rostral appendage and greenish turquoise extremities ( Fig. 2 View FIGURE 2 A). The colour of the legs is induced only by the coloured tubercle scales. The body is pale green or light brown with two dark brown spots and (occasionally) a beige lateral stripe on each side that stops at the base of the tail. The tail is the same colour as the body and (in stress colouration) possesses black annulations. The head is also greenish or brown with a dark stripe from the snout crossing the eyes to the occipital lobes. The skin around the mouth and the throat can be white. The colour description is based on a total of seven pictures of the Montagne d’Ambre form, referred to as C. boettgeri in Schmidt et al. (2010), C. boettgeri (picture 1c) in Glaw & Vences (2007) Calumma sp. in Nečas (2004) and C. boettgeri in Garbutt et al. (2001). The body and head of females is brown; the rostral appendage can be coloured bright blue, see picture of C. boettgeri in Schmidt et al. (2010).
The body colouration of males from Nosy Be in contrast is yellowish or greenish brown with little dark brown rosettes, when stressed ( Fig. 2 View FIGURE 2 B). The legs are brown with little blue or green spots resulting from the tubercles. The colouration of the head is similar to the body colouration. The rostral appendage differs clearly from Montagne d’Ambre with the absence of any striking colour and is the same brown colour as the casque. Females are uniformly light or greenish brown coloured. Compare also a total of seven pictures of Calumma boettgeri in Hyde Roberts & Daly (2014), in Glaw & Vences (2007: 191) picture 1a and 1b, in Nečas (2004) and in Henkel & Schmidt (1995).
Osteology of the skull based on micro-CT scans. Micro-CT scans of heads of two males and two females from Nosy Be and from Montagne d’Ambre exposed additional differences between the two forms. Specimens from Montagne d’Ambre ( Fig. 4 View FIGURE 4 , D and E) bear tubercles on the parietal in both sexes. These form a little parietal crest in the middle with three to four tubercles, laterally followed by two tubercles on each side. The frontal is also irregularly spotted with tubercles. The parietal and frontal of animals from Nosy Be in contrast are smooth ( Fig. 4 View FIGURE 4 , A and B).
As in all species of the genus Calumma , the nasal bones are paired ( Rieppel & Crumly 1997). These are broader in our specimens from Montagne d’Ambre (mean NW 0.35 mm vs. 0.24 mm in Nosy Be; mean NW/NL 0.18 vs. 0.14, Table 3 View TABLE 3 , Fig. 4 View FIGURE 4 ) and the anterior tip of the frontal bone does not exceed more than a half of the naris. In skulls from Nosy Be it does exceed this point, and the frontal meets the premaxilla, as described for C. nasutum ( Rieppel & Crumly 1997) . The parietal also varies between the two localities. In Nosy Be samples, the parietal tapers more tightly. Its diameter is at the tightest area on average 0.61 mm (vs. 1.06 mm) and 11% of the largest diameter of the parietal (vs. 22%, Table 3 View TABLE 3 , Fig. 4 View FIGURE 4 ). The parietal in Montagne d’Ambre samples appears wider and more compact. However, the form of the parietal is variable within localities and cannot be used as a diagnostic character. Although chameleons are sexually dimorphic animals, differences between sexes in skull structure were not proven ( Table 3 View TABLE 3 ).
Notes: m, male; f, female; NL, nasal length; NW, nasal width; RNWL, ratio of nasal width to length; PL, largest diameter; PS, parietal smallest diameter; RPSL, ratio of parietal smallest to largest diameter; PC, parietal crest absent (-) or number of tubercles.
Hemipenial morphology based on micro-CT scans. The scans of hemipenes of specimens from each population enable a detailed view of their structure. The hemipenes are illustrated in sulcal and asulcal view with the apex on top ( Fig. 5 View FIGURE 5 ). Both populations show large and deep calyces with smooth ridges on the asulcal side of the truncus. The apex is ornamented with two pairs of long pointed papillae and two pairs of rotulae. The papillae rise from the sulcal side of the apex and are curved to the asulcal side. They can be completely everted ( Fig. 5 View FIGURE 5 , C) or retracted in the apex ( Fig. 5 View FIGURE 5 , D). One pair of rotulae is placed on the asulcal side (the smaller one) and one pair on the sulcal side. Here some differences between the populations are recognizable; in Nosy Be (n = 2) the rotulae are slightly more denticulated, with 6–11 tips on asulcal side and 14–16 tips on sulcal side, compared to Montagne d’Ambre (n = 3) with 6–8 tips on the rotulae of the asulcal side and 11–14 tips on both rotulae on the sulcal side.


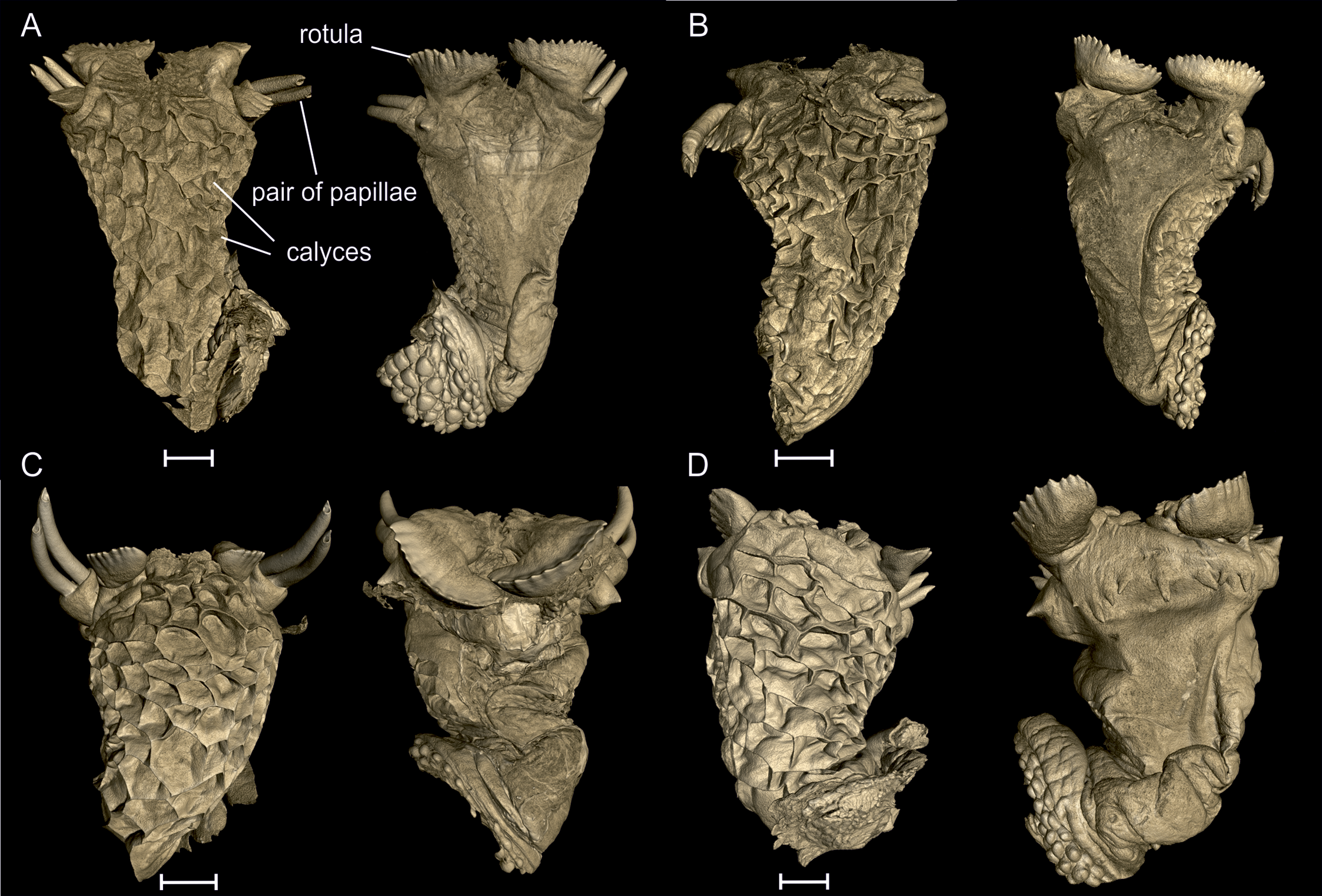

TABLE 2. Mean values and standard deviations (SD) of morphological measurements of Calumma boettgeri, n (males) = 11, n (females) = 12 and C. linotum males (n = 5); all measurements in mm. For abbreviations see Table 1.
| mean value SD | mean value | SD | mean value | SD | |
|---|---|---|---|---|---|
| SVL | 48.5 4.2 | 48.6 | 3.8 | 51.6 | 9.0 |
| TaL | 50.8 4.2 | 47.7 | 3.7 | 56.2 | 10.8 |
| TL | 99.3 8.1 | 96.3 | 7.2 | 107.8 | 19.7 |
| RST | 0.96 0.04 | 1.02 | 0.04 | 0.92 | 0.03 |
| LRA | 3.1 0.5 | 2.9 | 0.9 | 4.1 | 0.9 |
| RRS | 0.065 0.014 | 0.059 | 0.018 | 0.078 | 0.008 |
| RAPSC | 18.9 4.7 | 16.4 | 3.4 | 15.5 | 2.1 |
| NPSCM | 6.1 1.0 | 6.3 | 1.5 | 4.2 | 1.3 |
| NSL | 12 0.5 | 12 | 1 | 13 | 0 |
| NIL | 11.8 0.8 | 11.9 | 0.9 | 12.5 | 0.6 |
| OLD | 3.8 0.6 | 3.7 | 0.4 | 4.0 | 0.7 |
| DSC | 0.36 0.08 | 0.32 | 0.05 | 0.64 | 0.15 |
| RSB | 0.0075 0.0015 | 0.0066 | 0.0012 | 0.0124 | 0.0019 |
| NSC | 9.9 2.0 | 10.6 | 1.8 | 20.2 | 1.5 |
| UAD | 2.1 0.3 | 2.0 | 0.2 | 2.7 | 0.6 |
| RAS | 0.042 0.005 | 0.041 | 0.004 | 0.053 | 0.004 |
TABLE 3. Osteological measurements of important characters of the skull for differentiation between Calumma boettgeri und C. linotum.
| collection no. | species | locality | sex | NL | NW | RNWL | PL | PS | RPSL | PC |
|---|---|---|---|---|---|---|---|---|---|---|
| ZSM 440/2000 | C. boettgeri | Nosy Be | m | 2.1 | 0.3 | 0.14 | 4.7 | 0.5 | 0.11 | - |
| ZSM 441/2000 | C. boettgeri | Nosy Be | f | 2.2 | 0.2 | 0.09 | 3.9 | 0.5 | 0.13 | - |
| ZSM 444/2000 | C. boettgeri | Nosy Be | m | 1.8 | 0.3 | 0.17 | 4.7 | 0.3 | 0.07 | - |
| ZSM 227/2002 | C. boettgeri | Nosy Be | f | 1.3 | 0.2 | 0.15 | 4.1 | 0.6 | 0.15 | - |
| mean value | 1.85 | 0.25 | 0.14 | 4.35 | 0.49 | 0.11 | ||||
| SD | 0.40 | 0.06 | 0.03 | 0.41 | 0.11 | 0.03 | ||||
| MCZ 5988 | Ch. macrorhinus | ‘Madagascar’ | f | 2.0 | 0.2 | 0.10 | 3.9 | 1.1 | 0.28 | 1 |
| ZSM 2072/2007 | C. linotum | M. d'Ambre | m | 2.5 | 0.4 | 0.16 | 4.5 | 1.5 | 0.33 | 4 |
| ZSM 2073/2007 | C. linotum | M. d'Ambre | m | 2.1 | 0.4 | 0.19 | 4.6 | 0.6 | 0.13 | 3 |
| ZSM 873/1920/2 | C. linotum | M. d'Ambre | f | 2.3 | 0.3 | 0.15 | 4.2 | 0.9 | 0.21 | 4 |
| ZSM 551/2001 | C. linotum | Andampy | f | 1.4 | 0.3 | 0.23 | 4.3 | 0.9 | 0.21 | 3 |
| mean value | 2.07 | 0.36 | 0.18 | 4.40 | 0.97 | 0.22 | 3.50 | |||
| SD | 0.49 | 0.04 | 0.04 | 0.18 | 0.38 | 0.08 | 0.58 | |||
| ZSM 21/1923 | C. linotum | ‘Madagascar’ | m | 2.7 | 0.3 | 0.11 | 4.6 | 1.4 | 0.30 | 4 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.