
Dryomys nitedula (Pallas, 1778)
|
publication ID |
https://doi.org/ 10.5281/zenodo.6604339 |
|
DOI |
https://doi.org/10.5281/zenodo.6604292 |
|
persistent identifier |
https://treatment.plazi.org/id/9B215C43-FFD0-DD16-C972-F7D8FBA5FA11 |
|
treatment provided by |
Carolina |
|
scientific name |
Dryomys nitedula |
| status |
|
24. View On
Eurasian Forest Dormouse
French: Lérotin de forét / German: Baumschlafer / Spanish: Liron euroasiatico
Other common names: Forest Dormouse
Taxonomy. Mus nitedula Pallas, 1779 ,
Kazan region, near Volga River, Republic of Tatarstan.
Records from Balochistan, southern Pakistan, have not been verified; see comments under D. niethammeri . Confusion exists as to the publication date of the original description ofthis species by P. S. Pallas in part two of his Novae Species Quadrupedum e Glirum Ordine, but C. D. Sherborn in 1891 clarified the date to be 1779.
The type locality is often listed in the literature as “Lower Volga, Russia,” but as noted by S. I. Ognev in 1947, Pallas described this species based on specimens from oak forests of the Kazan region within what is now the Republic of Tatarstan. Pallas’s original 1779 text refers to “ad Volgam,” indicating the vicinity of the Volga River. Examination of museum specimens and detailed discussions of morphological variation and biogeography by G. Storch in 1978, B. Krystufek and V. Vohralik in 1994, M. G. Filippucci and colleagues in 1994, and others led M. E. Holden in 1993 and 2005 to surmise that more than one speciesis likely contained within D. nitedula . An allozymic and biometric study of southern populations by Filippucci and colleagues in 1994 provided strong support for recognizing the Israeli population of Dryomys as a valid species, a conclusion bolstered by ecological differences documented by E. Nevo and E. Amir in 1961, phallic and bacular morphology by S. Simson and colleagues in 1994, and multivariate and genetic analyses by N. Yigit and colleagues in 2011. Filippucci and colleagues in 1994 discussed the problem of applying an existing scientific name to the Israeli population. Results from the mtDNA phylogeographic study by O. O. Grigoryeva and colleagues in 2014 and 2015 revealed that the Caucasus population of Dryomys (formerly recognized as a subspecies, D. nitedula ognevi ) and central Russian Plain populations have a cytochrome-b gene sequence divergence value of 9-3% thatis indicative of species-level divergence. They hypothesized that the gap in forested steppe between the lower Don and Kuban rivers resulted in prolonged isolation of the Caucasus populations throughout the Pleistocene. Besides species level differences between the central Russian Plain and Caucasian populations, Grigoryeva and colleagues found that the Caucasian populations contains two sublineages: one occurs in the western Caucasus, and was described as a new subspecies, heptneri, and the other inhabits the central and eastern Caucasus. The cytochrome-b gene sequence divergence value between the two Caucasian lineages is ¢.6%, which may indicate that the two Caucasian populations represent two closely related species; Grigoryeva and colleagues cautioned that further study is needed to clarify the taxonomic status of these populations. Whether Caucasian populations represent one or more valid species, applying the correct existing scientific name is problematic. As discussed by Krystufek & Vohralik in 2005, geographicallimits of historically described subspecies and synonymy of subspecies names have not been documented. A comprehensive study that includes samples of all type specimens of all relevant named D. nitedula taxa is required to definitively assign the appropriate scientific name to the putative Caucasian and Israeli species. Monotypic.
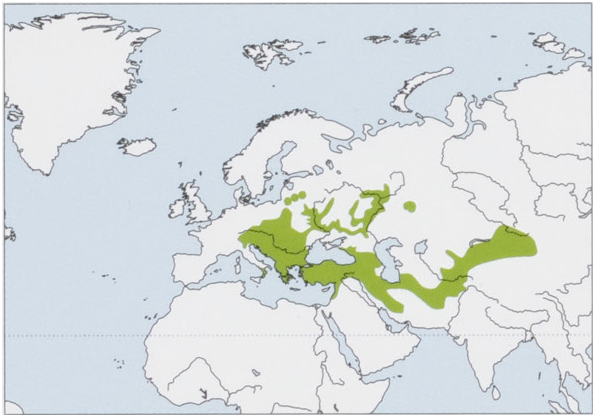
Distribution. From E Switzerland through E & S Europe, Anatolia and the Caucasus to C Russia and C Asia, reaching as far as 90° E in NW Xinjiang (China), and W Mongolia. View Figure
Descriptive notes. Head-body 79-103 mm, tail 66-96 mm, ear 11-15 mm, hindfoot 17-23-5 mm. In Lithuania, average body weight of adults is ¢.30 g in summer but c.43 g before hibernation. Tails of the Eurasian Forest Dormouse sometimes are truncated by damage because of false tail autotomy. Bushy tail gives it a squirrel-like appearance. Across the large distribution, dorsal pelage of adults varies from pale ash-gray to grayish brown and even bright rust, while under parts are yellowish white. Black stripes surround eyes and extend to rounded and short ears, forming conspicuous facemask. Tail is uniform in color but grayer than dorsum. Condylobasal length of skull is 21-25-3 mm, zygomatic breadth is 13-6-16-3 mm, and upper tooth row length is 3-5—4-6 mm. Angular process is perforated. External and cranial measurements for adults after first hibernation are from former Czechoslovakia and Austria. Eurasian Forest Dormice are larger and heavier in eastern part of the distribution, especially in Mongolia and China. Chromosome numberis stable at 2n = 48 throughout its distribution. Females have four pairs of nipples (1 pectoral + I abdominal + 2 inguinal = 8).
Habitat. Variety of habitats including broad-leaved, mixed, and coniferous forests; orchards; and Mediterranean evergreen shrubs. Eurasian Forest Dormice are widespread in mountains within different types of forest, including dwarf mountain pine ( Pinus mugo, Pinaceae ) woodland and rocky areas devoid of trees and shrubs. Presence of shrub layer and dense young trees in understory are main habitat requirements, and the common name “Bush Dormouse” would be more appropriate. In different parts ofits distribution, Eurasian Forest Dormice mainly use two types of nests: those situated in closed cavities in tree hollows or nest boxes, or nests constructed among thick and preferably thorny twigs.
Food and Feeding. Vegetation forms most of the diet of the Eurasian Forest Dormouse in Central Asia, the Caucasus, and Moldova where orchards are present or wild fruiting trees are abundant. They feed on apricots, apples, pears, plums, oak acorns, beechnuts, and also insects. Proportion of food of animal origin is higher in diets of Eurasian Forest Dormice in northern and western parts of the distribution, where they prey on birds that nest in nest boxes or tree hollows, consuming adult birds, fledglings, and eggs. Adult insects, insect larvae, and millipedes make up a significant proportion of their diet. Inflorescences of different trees, raspberries, bilberries,fruit of glossy buckthorn ( Frangula alnus, Rhamnaceae ), and hazel nuts also are eaten.
Breeding. Considerable variation exists in duration of breeding season, number of litters, and litter size across the distribution of the Eurasian Forest Dormouse. In Europe, only one litter is produced per year, and young are born in May-June. Average litter size varies from 3-2 young in Lithuania to 4-7 embryos in central Ukraine; four is the most frequent litter size recorded (range 1-6). In the Caucasus, Central Asia, Mongolia, and China, two litters may be produced per season. In Israel, breeding season occurs in March-December, and females may have been recorded to produce 2-3 litters/season, but average littersize of only 2-7 young (range 1-4) is the lowest across the distribution of the Eurasian Forest Dormouse. In central and eastern parts of the distribution, average litter sizes are comparatively high: 4-3 in Tajikistan, 4-6 in Kirgizstan, 5-1 in Azerbaijan, and 5-7 (range 3-9) in Mongolia. Gestation lasts 27-28 days. At c.1-5 months old, young can live independently. Females become reproductively active after their first hibernation. Six years is the maximum longevity recorded in the wild, but one individual survived more than eight years in captivity.
Activity patterns. The Eurasian Forest Dormouse is predominantly nocturnal but also crepuscular. In Romania, activity begins just before sunset in summer but about onehalf hour after sunset in autumn; it usually ends about half-hour before sunrise, independent of season. Daytime activity has also been recorded, and it is supposed to be associated with mating season. Daily torpor is infrequent in free-ranging Eurasian Forest Dormice. Activity season lasts only c¢.4-5 months from late April through early September on northern edge of the distribution (e.g. Lithuania and Tatarstan), but it is reportedly longer in southern populations. For example, they are active for up to eight months in Bulgaria (April-November). Like most other dormouse species, Eurasian Forest Dormice hibernate during the rest of the year, but winter activity was recorded in Bialowieza Forest, Poland, during comparatively warm periods. Hibernation nests are situated underground, often undertree roots. In the south-western part of its distribution in Israel, they are active throughout the year with short periods of torpor in winter.
Movements, Home range and Social organization. Eurasian Forest Dormice are arboreal and predominantly solitary. They are excellent climbers, but they also descend to the ground. They are usually solitary, but pairs of adult males and females are observed in the breeding season. Cohabitation of two adult females in the same shelter without communal nesting has occasionally been recorded. Groups of different age composition are most often found in nest boxes when juveniles become independent. Home ranges of adults partly overlap those of several neighboring dormice. Home ranges of males are larger than those of females. Average densities are comparatively very low in Eastern Europe; less than 1 ind/ha has been recorded in Lithuania, Poland, and Ukraine, but densities up to 20 ind/ha were estimated in Moldova, Armenia, and Mongolia. Eurasian Forest Dormice produce many high-frequency vocalizations, including ultrasound signals that appear to be social.
Status and Conservation. Classified as Least Concern on The [UCN Red List. The Eurasian Forest Dormouse has a wide distribution and stable population trend throughout most of its distribution. They have broad habitat tolerances and are found in many protected areas. In Europe, they are considered rare and are protected by international law under the European Union Habitats and Species Directive and the Bern Convention where these apply. The Eurasian Forest Dormouse is red-listed in some European countries, mainly those situated at the periphery of its distribution (e.g. Germany, Latvia, Lithuania, Poland, Romania and Switzerland, and also Mongolia). Despite its fragmented distribution, the Eurasian Forest Dormouse has the widest distribution of all Palearctic dormouse species.
Bibliography. Airapetyants (1983), Andera (1987 2011), Angermann (1963), Arslan & Zima (2014), Batsaikhan et al. (2008), Corbet (1978), Davydov (1984), Dogramaci & Kefelioglu (1990), Duma & Giurgiu (2012), Ellerman & Morrison-Scott (1951), Filippucci et al. (1994), Golodushko & Padutov (1961), Graphodatsky (2006), Grigoryeva, Balakirev, Stakheev et al. (2014), Grigoryeva, Balakirev, Sycheva et al. (2015), Grigoryeva, Krivonogov et al. (2015), Habibi (2004), Hassinger (1973), Holden (1993, 1996a, 2005), Juskaitis (1999, 2015), Juskaitis & Baltrunaite (2013b), Juskaitis et al. (2012), KryStufek (1985, 1999b), Krystufek & Vohralik (1994, 2005), Likhachev (1972), Lozan (1970), Lozan et al. (1990), Markov et al. (2009), Mitsainas et al. (2008), Mohammadi et al. (2013), Moradi-Gharahkloo (2009), Nevo & Amir (1961, 1964), Nowakowski (2001), Nowakowski & Godlewska (2006), Ognev (1947), Pallas (1779), Peshev & Delov (1994), Pilats et al. (2012), Roberts (1977 1997 2005), Rossolimo et al. (2001), Samarskii & Samarskii (1979), Satunin (1920), Scinski & Borowski (2006), Sekeroglu & Sekeroglu (2011), Sherborn (1891), Simson et al. (1994), Smith (2013), Spitzenberger & Bauer (2001a), Srinivasulu & Srinivasulu (2012), Storch (1978), Stubbe, Dawaa & Dorz (1986), Stubbe, Stubbe et al. (2012), Yigit, Colak, Colak, Ozkan & Ozkurt (2003), Yigit, Colak, Colak, Ozliik et al. (2011), Zima et al. (1994).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.