
Cristarmadillidium cabanillasi, Garcia, 2023
|
publication ID |
https://doi.org/10.11646/zootaxa.5323.2.8 |
|
publication LSID |
lsid:zoobank.org:pub:D986DFA4-F3EF-4E38-80B6-5FAA10D317C6 |
|
DOI |
https://doi.org/10.5281/zenodo.8204298 |
|
persistent identifier |
https://treatment.plazi.org/id/6236E7C0-6334-4091-9893-10C49459F805 |
|
taxon LSID |
lsid:zoobank.org:act:6236E7C0-6334-4091-9893-10C49459F805 |
|
treatment provided by |
Plazi |
|
scientific name |
Cristarmadillidium cabanillasi |
| status |
sp. nov. |
Cristarmadillidium cabanillasi View in CoL sp. nov.
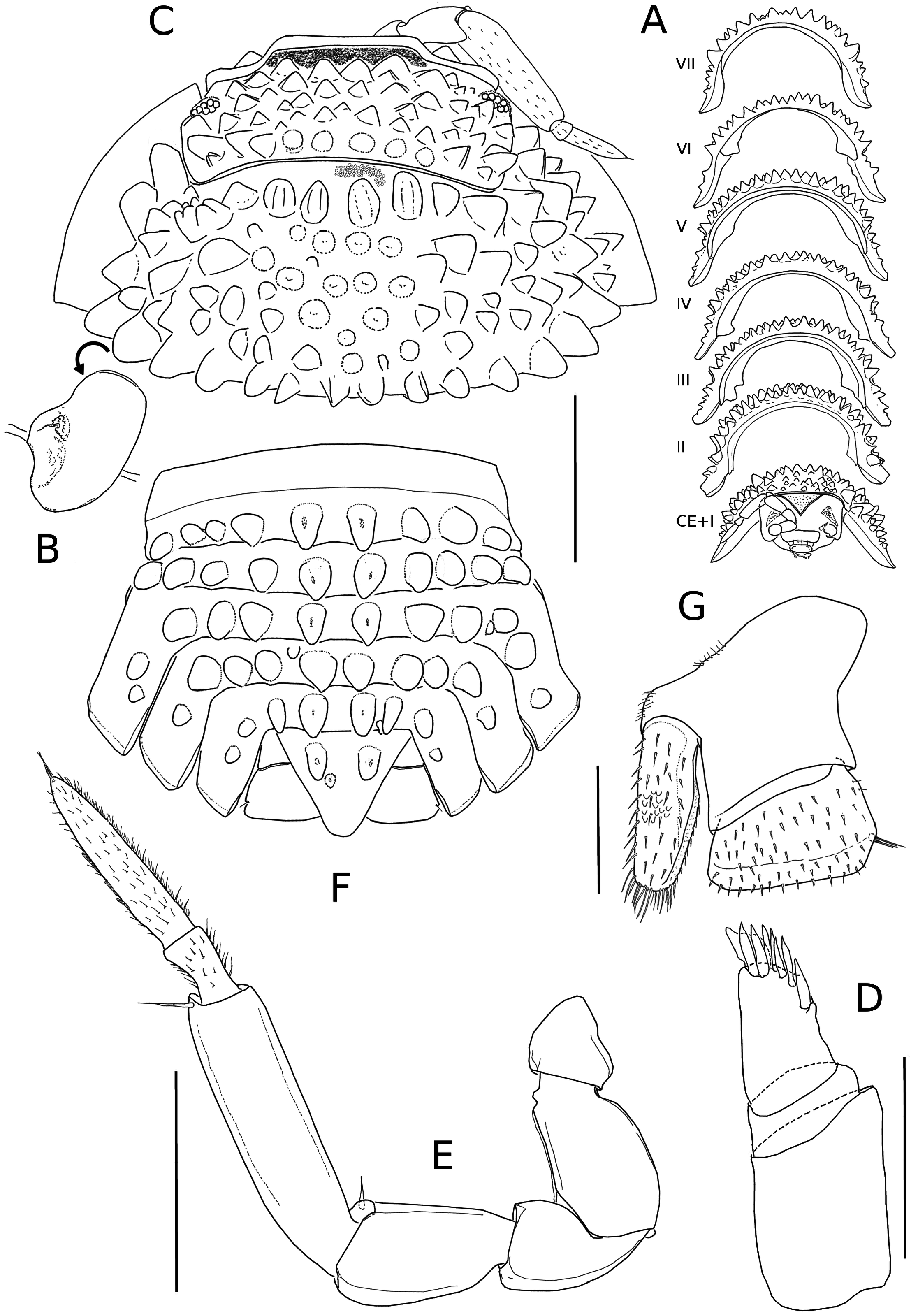
Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4
urn:lsid:zoobank.org:act:
Material examined. Holotype: ♀, Cadalso de los Vidrios, Madrid, Spain, leg. David Cabanillas-Roldán, 20–XII– 2020 ( MBCN 26801 ) . Paratypes: 2 ♁, 5 ♀, same locality as the Holotype, leg. David Cabanillas-Roldán, 20–XII– 2020 ( MNCN 20.04 About MNCN /20710 to 20.04/20716) . 1 ♁ (body parts of the dissected specimen, mounted in slides or stored in vials, as appropriate), same locality as the Holotype, leg. David Cabanillas-Roldán, 20–XII–2020, ( CLLG 1071 ) . 2 ♀, Villanueva del Pardillo , Madrid, Spain, leg. David Cabanillas-Roldán, 1–XI–2020 ( MBCN 26802–26803 ) . 4 ♁ ( MBCN 26804 to 26807 ) , 2 ♀ ( MBCN 26808–26809 ) , 1 immature ( MBCN 26810 ) , same locality as the preceding, leg. David Cabanillas-Roldán, 25–XI–2020 .
Diagnosis: Cephalon Eluma - type, with strong linea frontalis, without post-scutellar ridge; frontal shield separated from vertex and protruding over it. Epimera slightly extended laterally. Back with hypertrophic triangular tubercles; first pereon tergite with six transversal rows of tubercles; pereon tergites 2–7 with two main transversal rows of tubercles; pleon and pleotelson with one row of tubercles; posterior row of pereon-tergites with 14 regularly aligned tubercles; rows of pleonites 1–4 with 10 tubercles each; pleonite 5 with 8 tubercles; pleotelson triangular, as wide as long, with rounded tip and 2 dorsal tubercles.
Description: Maximum body length: male, 7.2 x 3.2 mm; female, 9 x 4 mm.
Habitus and general body form, as in Figs 1A–B View FIGURE 1 and 2 C–F View FIGURE 2 .Animals with perfect endo-conglobation (eusphaeric conglobation) ( Fig. 1A View FIGURE 1 ).
Colour light brown; edges of epimera, antennae, pereopods and pleopods without pigments ( Fig. 1A, B View FIGURE 1 ).
Body ( Figs 1B View FIGURE 1 , 2A, C, F View FIGURE 2 ) convex with epimera slightly extended laterally, more on pleon than on pereon. Dorsum with strong triangular tubercles compressed laterally; scale-setae triangular, tricorn-type, with rounded tip; integument covered with tiny round raised papillae. Noduli laterales ( Fig. 2B View FIGURE 2 ) small ( 0.03–0.05 mm), located medially on concave outer face of second tubercle (posterior row) of pereonites 1–7, not visible in dorsal view. No visible glandular pores. Dorsal tubercles arranged as follows ( Fig. 2A, C, F View FIGURE 2 ): cephalothorax with four rows of tubercles irregularly distributed, except rear row, consisting of 14 tubercles, larger than anteriors; pereon-tergite 1 with six rows of tubercles; pereon-tergites 2–7 with two rows and some smaller tubercles between them; posterior row of all pereon-tergites with 14 aligned tubercles; pleon-tergites 1–4 with one row of ten tubercles; pleon-tergite 5 with one row of eight tubercles; pleotelson with two tubercles. Cephalothorax ( Fig. 2A, C View FIGURE 2 ) Eluma - type, without postscutellar line; scutellar ridge delimitied by linea frontalis on upper margin, convex superior margin; frontal shield triangular, concave lateral margins and excavated anterior surface, distinctly separated from vertex and protruding over it; antennary lobes well developed, obliques and directed frontwards; eyes small, with 6–10 ommatidia. First pereon-tergite ( Fig. 2 C View FIGURE 2 ) without schisma, with angled posterolateral edge and acute anterior tip, extending beyond eyes position. Pleotelson triangular, as wide as long, with rounded tip ( Fig. 2F View FIGURE 2 ).
First antenna ( Fig. 2D View FIGURE 2 ) basal article longest, medial article short, distal article with 7–8 aesthetascs and triangular lobe. Second antenna ( Fig. 2E View FIGURE 2 ) short, just reaching first pereon tergite posterior margin; distal article of flagellum 2.7 times longer than basal.
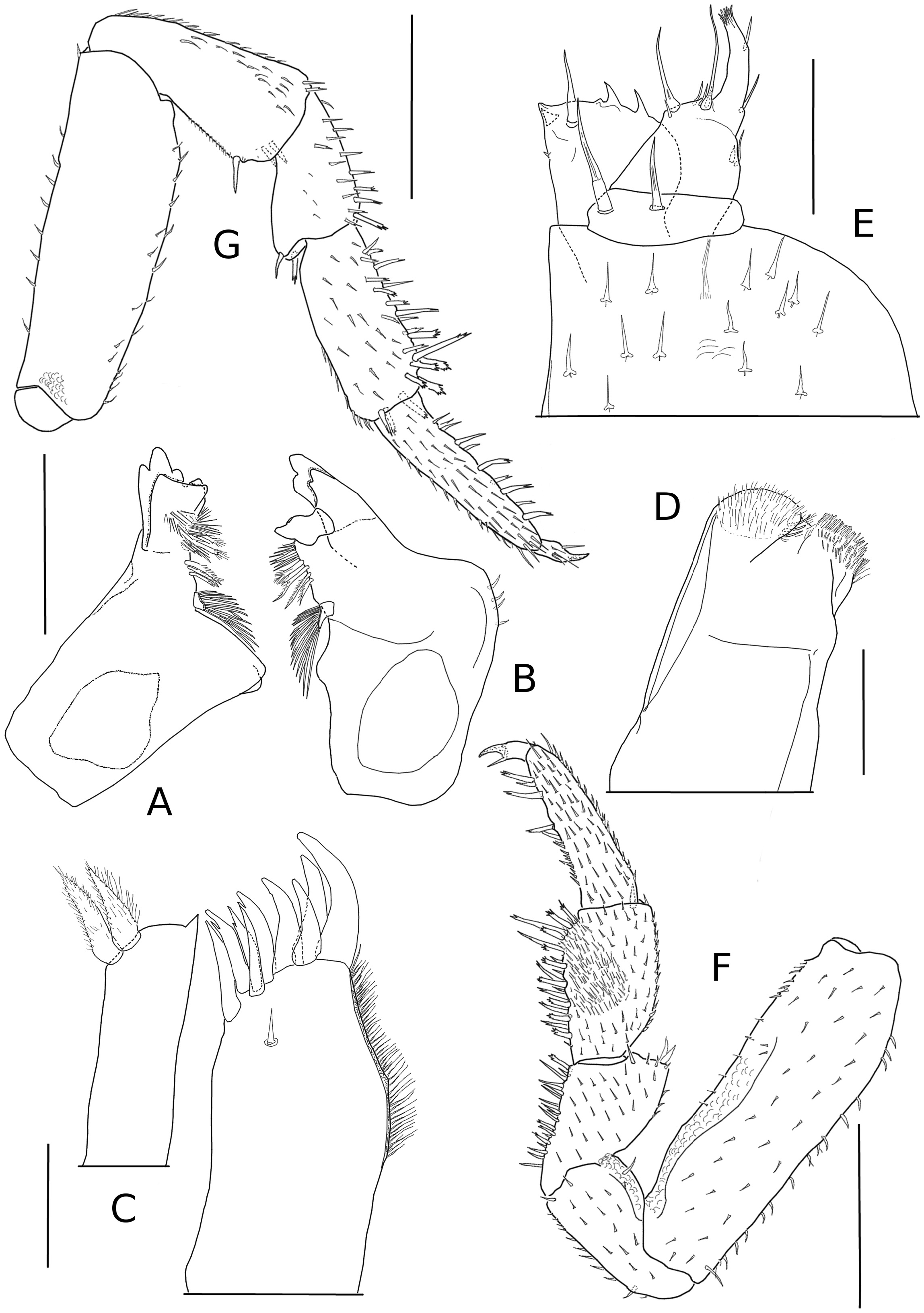
Mandibles with dichotomized molar process, about 15 branches; right mandible ( Fig. 3A View FIGURE 3 ) with 2 + 3 penicils; left mandible: ( Fig. 3B View FIGURE 3 ) 1 + 3 penicils. Maxillule ( Fig. 3C View FIGURE 3 ) external branch with 4 + 5 teeth, three of them cleft at apex; internal branch with two thick penicils and wide triangular posterior corner. Maxilla (Fig, 3D) with subequal lobes, external lobe bearing three thick setae. Maxilliped ( Fig. 3E View FIGURE 3 ) palp with two setae on first article, second article with two long setae and few small setae, distal article with tuft of short setae and two long setae on external margin; endite with rounded external margin, two triangular teeth on anterior edge and strong medial seta on caudal face.
Pereopods with more ventral setae in males than in females; dactyli with inner claw shorter than outer one. Pereopod 1 with oblique antennal grooming brush.
Pleopods 1 and 2 exopods with monospiracular pleopodal lungs.
Uropods with trapezoidal, plate-like, exopod about twice wide than long; endopod inserted proximally on medial margin of protopod, not surpassing the posterior margin of exopod.
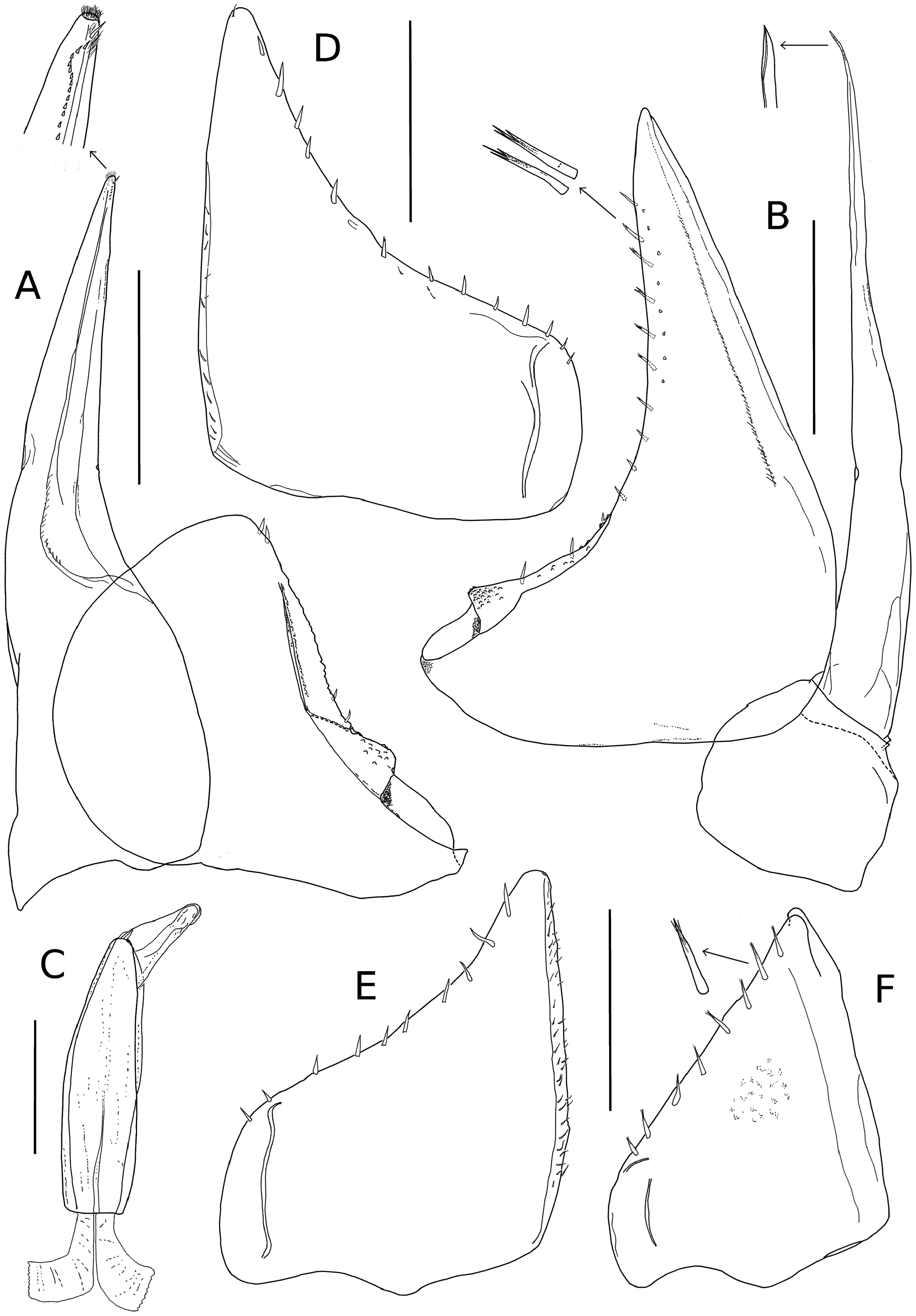
Male. Pereopods 1–6 ( Fig. 3F View FIGURE 3 ) with ventral brush of setae on meropod and carpopod. Pereopod 7 ( Fig. 3 G View FIGURE 3 ) ischiopod with straight ventral margin. Genital papilla as in Fig. 4C View FIGURE 4 . Pleopod 1 ( Fig. 4 A View FIGURE 4 ) exopod wider than long; posterior lobe subtriangular with rounded tip; respiratory field oblique, with protruding edge on its ventral face partially covering spiraculum; endopod straight, twice as long as exopod. Pleopod 2 ( Fig. 4 B View FIGURE 4 ) exopod triangular, longer than wide, bearing posterior lobe; respiratory field oblique, with protruding edge on its ventral face partially covering spiraculum; endopod flagelliform, longer than exopod. Pleopods 3–5 as in Fig. 4 D–F View FIGURE 4 .
Etymology: The new species is named after David Cabanillas-Roldán, a myriapodologist from Madrid ( Spain), who collected the material studied here.
Remarks: According to the original diagnosis of Cristarmadillidium ( Arcangeli 1936; Vandel 1954), C. cabanillasi sp. nov. can be accommodated in this genus because it presents the following morphological features: Eluma - type cephalon; eyes reduced; a lack of schisma on the first pereon-tergite, pereon-epimera 6–7 and pleon-epimera 3,4 and 5 expanded horizontally, and male pereopod 7 not specially modified. Recuero et al. (2021), based on morphological and molecular data, amend the diagnosis by adding, among other characteristics, the presence of a small schisma on the posterior edge of the first pereonite (only in C. myrmecophylum Recuero, Rodríguez-Flores & García-Paris, 2021 ); external margin of pereon-tergite 2 epimera straight; and distal part of pereonite 4-5 epimera narrow and rounded. (see discussion).
In having the dorsal surface tuberculate, C. cabanillasi sp. nov. is similar to C. muricatum (Budde-Lund, 1885) , type species of the genus; however, it differs in having the tubercles numerous, and less developed. Also, it differs from C. muricatum by the following characteristics: pereon and pleon-epimera less expanded laterally; integument with round raised papillae (diameter = 0.02 mm), instead of larger oval cavities (diameters = 0.04 x 0.06 mm); pleotelson triangular with straight sides and broadly rounded tip, instead of slightly concave lateral edges and pointed tip; anterolateral tip of first pereon-tergite triangular and acute, forming an angle with the anterior edge, instead of less prominent and rounded, without angular transition to anterior edge; epimera of pereon-tergite 5 broad and rounded instead of narrow and almost triangular; exopod of male first pleopod 1.2 times as wide as long instead of 1.4 times as wide as long; and uropod exopods ( Fig. 2 G View FIGURE 2 ) wider than long (ratio w/l=1.56) instead of as wide as long (ratio w/l=1), among other characteristics. The morphological differences with the remaining Cristarmadillidium species are even greater. From C. breuili Vandel, 1954 , and C. alticola Cifuentes, 2021 , it differs mainly by the hypertrophic tubercles on the body. From C. zaragozai Cifuentes & Prieto, 2020 , it can be distinguished mainly by the shape and structure of the tergal tubercles, which are well differentiated and raised in C. cabanillasi sp. nov. and most flattened, longitudinally keeled, and with a different arrangement in C. zaragozai . All of these species differ from C. myrmecophylum by lacking schisma on the first pereonite.
Cristarmadillidium cabanillasi sp. nov. also shows some morphological characteristics that bring it closer to the genera Estenarmadillidium Cifuentes, 2021 , and Iberiarmadillidium Recuero, Rodríguez-Flores & García-París, 2021 . These similarities are as follows: epimera of pereon-tergites less expanded outwards; integument covered with tiny round raised structures (papillae); lateral edge of the pereonite 5 straight with rounded angles and anterolateral tip of the first tergite extended forwards, forming an angle with the corresponding anterior edge ( Fig. 1 View FIGURE 1 ), among other features (see Cifuentes, 2021; Recuero et al. 2021). However, all species of these two genera have the first tergite with a schisma. Therefore, until a systematic review of these genera can be made, we believe it is convenient to keep the new species within the genus Cristarmadillidium .
Ecology: The specimens from the type locality, Villanueva del Pardillo, were collected from a pastured oak woodland. The specimens from Cadalso de los Vidrios were collected in a rocky clearing inside a forest of Pinus sylvestris L., under stones, with unidentified ants.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |