
Limnadopsis bloodwoodensis, Schwentner, Martin, Timms, Brian V. & Richter, Stefan, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.210812 |
|
DOI |
https://doi.org/10.5281/zenodo.5619654 |
|
persistent identifier |
https://treatment.plazi.org/id/9A5D7F26-FFD2-FFDF-7294-FAEFFA7FFE02 |
|
treatment provided by |
Plazi |
|
scientific name |
Limnadopsis bloodwoodensis |
| status |
sp. nov. |
Limnadopsis bloodwoodensis View in CoL sp. nov.
( Figs. 1 View FIGURE 1 a, b, 2, 3)
Limnadopsis sp. ‘Roskos’: Schwentner et al. 2011
Etymology: The name is derived from Bloodwood Station in northern New South Wales where the species was first found.
Type locality: New South Wales, Bloodwood Station, Roskos Paleolake, 29°27'42.9''S, 144°48'12.5''E, 19.ii.2010, collected by M. Schwentner and B.V. Timms
Type material: Holotype 3 AM P.85175 (GenBank HQ717790 View Materials ); allotype Ƥ AM P.87800 (GenBank JQ424941 View Materials ); paratype 1Ƥ AM P.85176 (GenBank HQ717791 View Materials )
Further material examined: Queensland: 13 (AM P.86421), 1Ƥ (AM P.86422), Wombah Station, Hatch Lake, 28°56'S, 144°54'E, 26.iii.2000; New South Wales: one resting egg retrieved from dry sediment sample (‘Egg51’; GenBank HQ717746 View Materials ), first pool East Mossgiel, 33°17'40.7''S, 144°42'54.3''E, 23.i.2010
Description.
Male (Holotype, AM P.85175) ( Figs. 1 View FIGURE 1 a, 2a, b)
Carapace. Length 16.8 mm, height 9.8 mm, length/height ratio 1.71 ( Fig. 1 View FIGURE 1 a). Ten growth lines, regularly arranged, with distance between growth lines increasing towards ventral margin, two additional growth lines crowded together at ventral margin; growth lines not distinct, surface between them smooth without structure, slightly translucent. Dorsal margin convexly rounded with concavity before umbo and no serration. Dorso-anterior corner about 130°, dorso-posterior corner protruding, posterior margin convex and extending beyond dorso-posterior corner, ventral margin evenly convex.
Head. Mound containing the eye protruding, frontal organ pyriform and situated halfway between mound and posterior end of head ( Fig. 2 View FIGURE 2 a). Rostrum extending at ~100° from head, moderately acute distally and slightly bent apically. First antenna (antennule) with twelve lobes ( Fig. 2 View FIGURE 2 a) reaching to first antennomere of second antenna. Peduncle of second antenna with ten discernable segments, each armed with a row of setae and spines; anterior ramus with 18 antennomeres (posterior ramus damaged), middle segments of both rami with six spines in row on anterior surface and eight long setae in row on posterior surface.
Trunk composed of 27 segments, eleven most anterior of which lack dorsal armature; next seven segments with several setae each, then eight segments with several dorsal spines each, and last segment without armature. Claspers and thoracopods typically spinicaudatan in structure ( Olesen et al. 1996; Timms 2009; descriptions herein based on paratype AM P. 86421). Clasper with protruding tumidity on proximal part of main hand ( Fig. 2 View FIGURE 2 e); third thoracopod with five endites and endopod (or endite 6) on medial margin and elongated exopod and oval epipod on lateral margin ( Fig. 2 View FIGURE 2 f); fifth endite not terminating in clearly discernable palp, equally thickened to tip with no obvious articulation.
Telson bearing 13 spines along each dorsal ridgeline ( Fig. 2 View FIGURE 2 b), first spine only slightly larger than following spines, these being equal in size and evenly spaced. Last spine enlarged with the two preceding spines originating at its base; large spine on right side of telson bent at approximately 90° to telson, that on left side at approximately 45°. Filaments inserted between third and fourth spines.
Furca. Each ramus bearing ten long setae on first quarter ( Fig. 2 View FIGURE 2 b), total of 13 spines on following two quarters. Spines decreasing in length and becoming stouter posteriorly, but last spine enlarged. Distal third of each ramus covered by small denticles. Proximal three-quarters of each ramus straight, distal quarter bent dorsally at approximately 45°. No spine or protrusion beneath the basal articulation.
Female (Allotype, AM P.87800) ( Figs. 1 View FIGURE 1 b, 2c, d)
Carapace. Length 16.3 mm, height 10.5 mm, length/height ratio 1.55 ( Fig. 1 View FIGURE 1 b). Nine growth lines, regularly arranged, with distance between growth lines increasing towards ventral margin, two additional growth lines crowded together at ventral margin; surface between them smooth and without structure, slightly translucent. Dorsal margin strongly convex with depression before umbo; dorsal serration weakly expressed on some growth lines. Dorso-anterior corner nearly a right angle, ventral margin slightly rounded, posterior margin convexly rounded and extending past dorso-posterior corner, latter a right angle.
Head. Mound containing the eye round and protruding; rostrum short, triangular ( Fig. 2 View FIGURE 2 c). First antenna with nine lobes reaching first antennomere of second antenna. Peduncle of second antenna with ten discernable segments, each with row of setae and spines; anterior ramus of second antenna with 15 antennomeres, posterior ramus with 14 antennomeres.
Trunk composed of 27 segments, with dorsal armature starting from eighth segment: segments eight to eleven with single tiny spine, segments 12–19 with numerous long, thin setae, segments 20–26 with short, stout spines, last segment without dorsal armature. Epipodites of all segments elongated, but longest and thickest on segments eight and nine.
Telson bearing twelve spines along each dorsal ridgeline (N.B.: the telson was damaged when this individual was still alive; it is likely that two spines are missing, and the crowding of the first four spines may be due to damage; Fig. 2 View FIGURE 2 d). First spine slightly enlarged, following spines equal in size; first four spines more closely spaced than following spines; last spine enlarged with two preceding spines originating from its base; large spines on both sides of telson bent at approximately right angle to telson.
Furca. Each ramus bearing ten setae followed by six spines, with last spine enlarged ( Fig. 2 View FIGURE 2 d); distal third of each ramus covered in denticles; whole ramus slightly bent, distal third bent dorsally at nearly right angles.
Eggs. Height and diameter about 195 µm ( Fig. 3 View FIGURE 3 a). Not spherical but resembling subconical cylinders with convex ends bearing irregular grooves. Sides of eggs are encircled by a series of long, slightly curved, horizontal grooves.
Variability. The variability of L. bloodwoodensis sp. nov. cannot be fully assessed as only five specimens are known. The carapace displays nine to twelve indistinct growth lines. Up to two additional growth lines were crowded together at the ventral margin in some specimens. Carapace is always slightly translucent with no sculpturing between growth lines. Dorsal margin can be straight or strongly convex. Females more often have a convex dorsal margin, though that of males can also be convex. Three specimens had minor serration at the dorsal margin of a few of outer growth lines. Posterior margin extends past the dorso-posterior corner.
All specimens studied had 27 trunk segments. Last segment always lacks dorsal armature ( Fig. 3 View FIGURE 3 d). It is preceded by six to nine spine-bearing and five to eight setae-bearing segments. First antenna has 7–13 lobes ( Fig. 3 View FIGURE 3 b) and reaches at most to the second antennomere of the second antenna. Second antenna has 14–17 antennomeres. Telson bears 12–15 evenly sized and spaced spines ( Fig. 3 View FIGURE 3 c). First spine is only slightly larger than the following spines; last spine is greatly enlarged with two to three smaller spines originating at its base. In females last spines on both sides of the telson are bent at approximately 90°, in males only the spine on the right side of the telson is bent, the other is nearly straight. Each furcal ramus has 8–13 setae. Several of the setae are thin without setules and resemble elongated spines. They are followed by 6–15 spines (last spine always enlarged) and a series of denticles. The scales on the tip of the movable finger of the male clasper are bifid, with the ventral part slender and palp-like and the dorsal part slender, weakly bent towards the movable finger and slightly serrated ( Schwentner et al. 2011). In all specimens studied only one single scale was present on each movable finger.
The genetic diversity observed in L. bloodwoodensis sp. nov. is low at only 0.0–1.0 % uncorrected p-distance and K2P corrected distance for COI (based on four specimens).
Distribution. Adults of this species have only been collected from two temporary pools in the Paroo River catchment on the border between New South Wales and Queensland. The occurrence of a resting egg in central New South Wales suggests a larger distribution range, at least within the Murray-Darling Basin.
Remarks. L. bloodwoodensis sp. nov. is similar to L. parvispinus Henry 1924 and L. occidentalis Timms 2009 in overall morphology, but there are important differences ( Table 1 View TABLE 1 ). The number of trunk segments is the most obvious, as both L. parvispinus and L. occidentalis have fewer than 27 segments. Furthermore, L. parvispinus has more carapace growth lines, more telsonic spines and fewer furcal spines, whereas L. occidentalis exhibits fewer lobes on the first antenna and fewer furcal spines than L. bloodwoodensis sp. nov. ( Table 1 View TABLE 1 ). Schwentner et al. (2011) delineated these three species on the basis of differences in the scale at the tip of the movable finger of the male clasper (the scales also differed in size, not shown in Table 1 View TABLE 1 ). Misidentifications are most likely to concern L. parvispinus as these two species co-occur in the Murray-Darling Basin, though probably not in the same pools.
Both habitats where adult L. bloodwoodensis sp. nov. were collected are hyposaline. At the time of collection Hatch Lake had a salinity of 3.3 g /L and the previously recorded range for this lake is 0.6–19 g /L ( Timms 2008). The salinity of Roskos Paleolake was not recorded during the collection event, but the previously recorded salinity range for this lake is 1.3–8.6 g /L. It is likely that L. bloodwoodensis sp. nov. only lives at the lower end of the salinity range (probably below 5 g /L), but it appears to be the only species of Limnadopsis which is adapted to hyposaline water. Limnadopsis paradoxa Timms 2009 has been reported from a lake with varying salinities, but only when the water was fresh (Timms 2009).
Phylogenetic analyses placed L. bloodowoodensis sp. nov. in a sister group relationship to L. parvispinus and L. brevirostris ( Schwentner et al. 2011) . L. occidentalis was not part of the phylogenetic analysis but may be closely related to this group.
Specimens identified as ‘ Limnadopsis tatei ’ by Weeks et al. (2009; GenBank FJ44181, FJ44182) differed by just 2 % uncorrected p-distance for the COI gene ( Schwentner et al. 2011). These specimens may thus also be L. bloodwoodensis sp. nov., which would extend the range of L. bloodwoodensis sp. nov. into the Northern Territory.

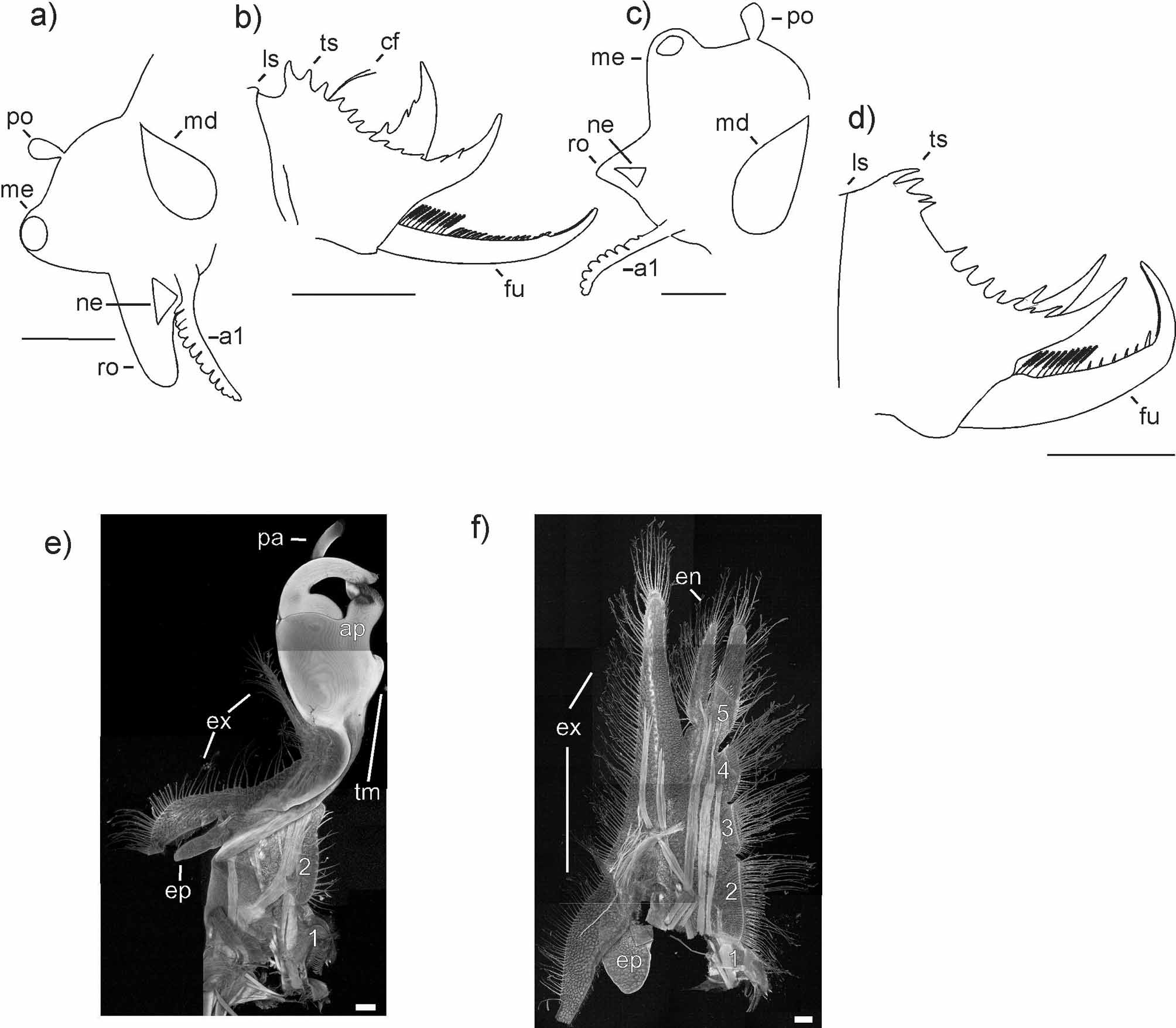

TABLE 1. Comparison of morphological characteristics of the species Limnadopsis parvispinus, L. occidentalis and L. bloodwoodensis sp. nov. The information on the first two species is taken from Timms (2009) and the information on the claspers from Schwentner et al. (2011).
| growth lines on carapace | L. parvispinus 10–18 | L. occidentalis 9–12 | L. bloodwoodensis sp. nov. 9–12 |
|---|---|---|---|
| trunk segments 1st antenna | 25–26 7–10 | 25 6–9 | 27 7–13 |
| 2nd antenna spines on telson | 14–21 16–22 | 15–16 12–16 | 14–17 12–15 |
| setae on furcal ramus spines on furcal ramus | 8–15 4–10 | 12 2–6 | 8–13 6–15 |
| eggs | subconical cylinders, base flat surface | round, short grooves | subconical cylinders, base convex surface |
| tip of movable finger of male clasper | 2–4 bifid scales | single bifid scale | single bifid scale |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |