
Dryopteris bernieri Tardieu
|
publication ID |
https://doi.org/ 10.5252/a2011n1a1 |
|
persistent identifier |
https://treatment.plazi.org/id/886CAA78-FFF0-FFE5-FC86-0EDFFBF6FD13 |
|
treatment provided by |
Carolina |
|
scientific name |
Dryopteris bernieri Tardieu |
| status |
|
2. Dryopteris bernieri Tardieu View in CoL ( Figs 8 View FIG ; 9 View FIG )
Notulae Systematicae (Paris) 15 (2): 161 (Apr. 1956). — Type: Bourbon ( Réunion), Lépervanche-Mézière 22 (holo-, P00349606!) .
OTHER MATERIAL EXAMINED. — Comoro Islands (Anjouan). Johanna Island, III.1977, Bewsher s.n. (K). Réunion. Bourbon, 1847, Armange s.n. (P00349616). — Bourbon, 1835, Bernier 96 (P00349608). — Route forestière du pic Maïdo, falaise ombragée d’une ravine, 1750 m, 2.XI.1973, Badré 773 (P00349619). — Palmiste rouge, vallée du Bras des Calumets, plantation d’ Eucalyptus, 1000 m , 15.XI.1973, Badré 902 (P00349617, P00349618). — Sentier de Cilaos vers l’îlet à Cordes, entre Cilaos et le bras Rouge, 17.XI.1973, Badré 974 (P00349620, P00349621). — Île Bourbon, 15.II.1847, Boivin s.n. (P00349609). — plaine des Cafres au pied du Piton de Villers, 7.VI.1851, Boivin s.n. (P00349612). — Oraire, VII.1851, Boivin s.n. (P00349611).— Île Bourbon, Boivin s.n. (P00349614). — Cilaos, prés de la chapelle, 600 m, 12.I.1963, Cadet 322 (P00349629). — Fissure profonde sommet du Rempart de Belle courbe, 2300 m, 12.IX.1968, Cadet 1542 (P00349628). — Haute vallée de la rivière des Remparts, sous-bois sur falaise, 1600 m, 20.XI.1968, Cadet 1713 (P00349626, P00349627). — La Réunion, sous-bois d’ Acacia et d’ Hypericum , bras Sec, cirque de Cilaos, 1300 m, 7.II.1969, Cadet 1904 (P00349624). — La Réunion, terrestre, sous-bois de végetation arbustive, cirque de Cilaos, 1200 m, 16.II.1973, Cadet 4115 (P00349622). — La Réunion, terrestre, sous- bois de forêt sèche, cirque de Grand Bassin, vers 1000 m, 28.VI.1973, Cadet 4375 (P00349623). — La Réunion, sous-bois de végétation arbustive pionnière, vallée du Bras Rouge, cirque de Cilaos au-dessous de l’Îlet du Salazes, 1300 m, 22.III.1974, Cadet 4559 (P00349625). — Île Bourbon, De Cordemoy s.n. (BM000801043). — Île Bourbon, VII.1837, Gaudichaud s.n. (P00349615-A only). — Réunion, Cilaos Distr., road from Cilaos to Îlet à Cordes on steep earthbank, in deep shade, occasional, 1000 m, 20.IX.1984, Jacobsen 5541 (FR). — Bourbon, Johnstone 80 (BM000801026). — Bourbon, Lépervanche-Mézière 22 (P00349606). — La Réunion, 1891, Maigre s.n. (P00349607). — Réunion, Potier s.n. (P00349610).
Sine Loco. Lépervanche-Mézière s.n. (P00349613).
DESCRIPTION
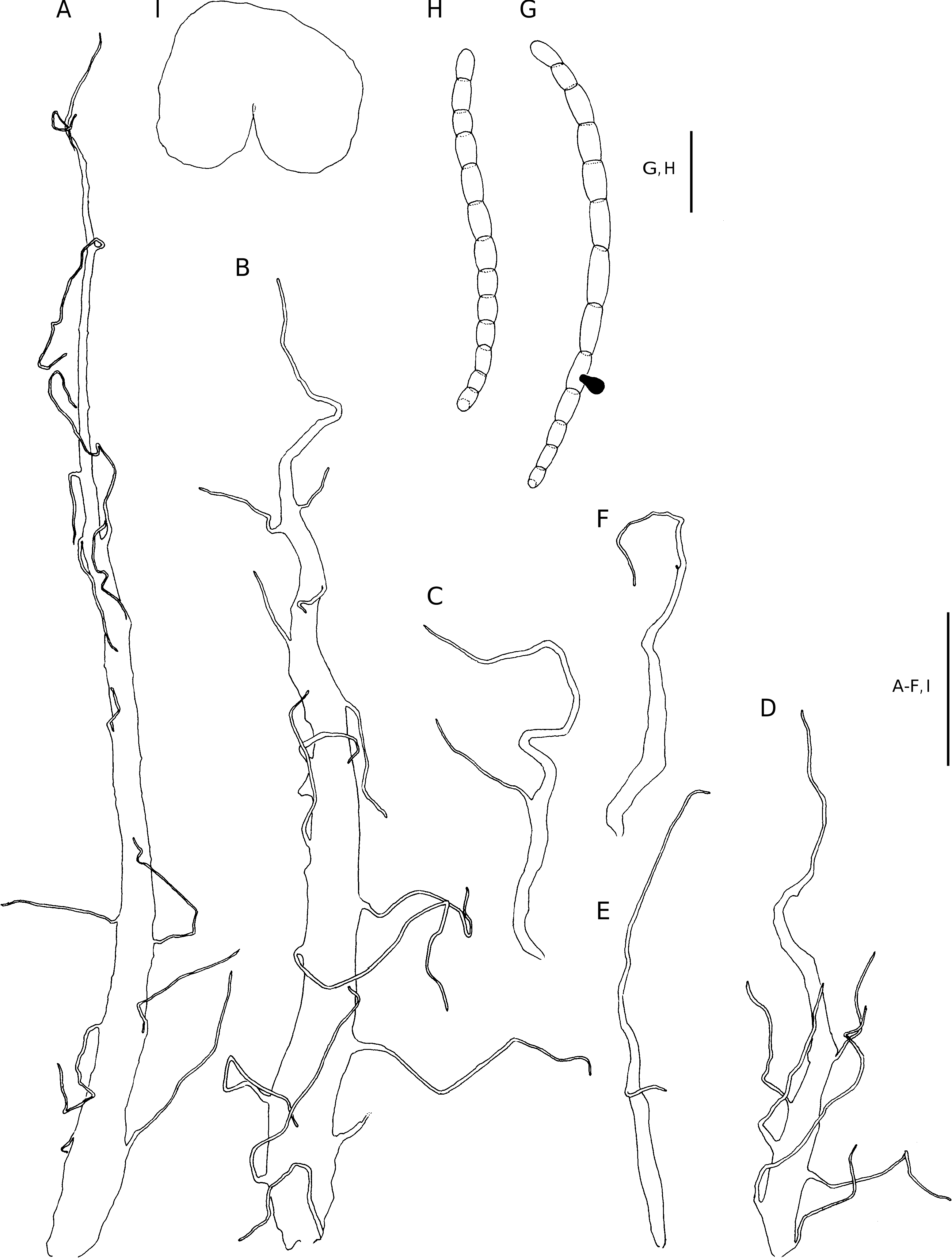
Plants terrestrial. Rhizome short-decumbent, up to 70 mm long and 16 mm in diameter, set with roots, closely spaced stipe bases and scales, the scales ferrugineous, chartaceous, adnate, linear-acuminate, up to 35 × 4.5 mm, regularly set with capitate glands along the margins and laminae, the apex terminates in a short series of oblong cells. Fronds crowded, arching, up to 1.2 m long; stipe up to 510 mm long and 7 mm in diameter, proximally castaneous, stramineous higher up, adaxially shallowly sulcate, proximally densely set with scales similar to those on the rhizome, up to 45 × 4 mm, moderately to closely set with capitate glands, sparsely scaled higher up, the scales ferrugineous to stramineous, sessile, ovatecaudate to narrowly lanceolate-caudate, up to 9 × 3 mm, cordate, the margins and laminae regularly set with capitate glands, often also with a few scattered pluricellular, mostly uniseriate hairs, which often is glandular, the scale apex terminates in a short series of oblong cells; lamina deltate to broadly ovate, up to 610 × 340 mm, to 2-pinnate-pinnatifid, pinnatifid towards the apex, with up to 8 petiolated pinna pairs; rachis ferrugineous, sulcate adaxially, narrowly winged towards the apex, closely glandular, sparsely scaled, the scales sessile, narrowly lanceolate-caudate to subulate-caudate, up to 4 × 0.6 mm, cuneate, the margins and laminae regularly set with capitate glands, the apex terminates in a short series of oblong cells; pinnae opposite to alternate, mostly overlapping, petiolate at base, adnate and basiscopically decurrent with rachis winged towards lamina apex, petiole up to 13 mm long, basal pair longest and conspicuously basiscopically developed, up to 1-pinnate-pinnatifid, inaequilaterally ovate, up to 290 × 150 mm, pinnae higher up more symmetric, oblong-acuminate to linear-acuminate, with up to 6 petiolated pinnule pairs; pinna-rachis sulcate adaxially, narrowly winged towards the apex, closely glandular, sparsely scaled abaxially, the scales similar, but smaller than those on the rachis; pinnules petiolate, the petiole up to 2.5 mm long, opposite to alternate, pinnatifid, not or slightly overlapping, firmly herbaceous, dark green adaxially, paler abaxially, pinnatifid, acroscopic pinnule on basal pinnae linear-acuminate to ovate, up to 70 × 24 mm, basiscopic pinnule on basal pinnae linear-acuminate to narrowly ovate,up to 90 × 32 mm, abaxially with isocytic hairs mostly along the veins, the hairs often with one or more glandular cells near the base; pinnule-rachis adaxially convex, narrowly winged, closely glandular adaxially and abaxially, the glands capitate, (60-)106(-140) µm long, and with scattered scales, the scales ferrugineous, chartaceous, narrowly lanceolate-caudate to subulate-caudate, glandular along the margins and laminae; segments proximally sessile, adnate higher up, oblong-obtuse to ovate obtuse, up to 20 × 8 mm, larger segments often shallowly lobed, denticulate, adaxially and abaxially closely set with capitate glands along and between the veins.Venation obscure adaxially,evident abaxially, lateral veins in segments simple or once forked, ending in the teeth near the margin.Stomata mostly of the polocytic type, (32-)47(-56) µm long. Sori essentially 2-seriate on pinnules, inframedial on predominantly anadromous vein branches, up to 1.5 mm in diameter and discrete at maturity; indusium castaneous, firmly herbaceous, up to 1.7 mm in diameter, reniform, often strongly recurved, entire to shallowly repand, eglandular or glandular along the margin, often also on the lamina. Sporangium stalk simple, with several hairs and/or glands, capsule with (12-)14(-17) indurated annulus cells, epistomium (3-)4(-7)-celled, hypostomium (4-)6-celled. Spores brown, (36-)42(-52) × (24-)29(-36) µm long.
DIAGNOSTIC FEATURES AND RELATIONSHIPS Dryopteris bernieri can be separated from other Dryopteris species from Réunion by the axes being closely set with capitate glands, a feature that is absent from other Dryopteris species on the island. Similar glands also occur adaxially and abaxially along and between the veins on the lamina. The scales of D. bernieri are also regularly set with capitate glands along the margins and laminae.
Fraser-Jenkins (1986) does not list the species, but it is here placed in section Marginatae Fraser- Jenk. The occurrence and distribution of capitate glands on the lamina and the similar scale morphology suggest it being related to D. glandulosopaleata J.P.Roux from Africa. Dryopteris bernieri is separated from D. glandulosopaleata in them being geographically isolated, and in the stomata and spores of D. bernieri being significantly smaller than that of D. glandulosopaleata .
DISTRIBUTION AND HABITAT
Dryopteris bernieri has a somewhat disjunct distribution occurring on Réunion, where it grows at elevations ranging between 600 and 2300 m on both Massif du Piton des Neiges and Massif de la Fournaise and on Anjouan (Johanna Island/ Anjouan), Comoros ( Fig. 10 View FIG ). Both massifs are the result of successive volcanic eruptions, but their subsequent erosion has resulted in the formation of radiating valleys with steep flanks. The species grows in deep shade in wet evergreen forests, on earth banks and screes along the steep sloped valleys, and in Eucalyptus plantations.Tardieu-Blot (1956a, 1958) noted that the species is also present in the “ Comores ” by citing a Boivin collection. I have not seen any collections of D. bernieri from that island group.
3. Dryopteris bojeri (Baker) Kuntze ( Figs 11 View FIG ; 12 View FIG )
Revisio generum plantarum 2: 812 (5 Nov. 1891). —
Nephrodium bojeri Baker , Synopsis filicum 7: 280 (4 Oct.
1867). — Type: Mauritius, Bojer s.n. (lecto-, K!, designated
by Roux [2009: 121]; isolecto-, M0065885!).
Nephrodium amplum Bory, Voyage aux Indes Orientales. Botanique 2 (1): 62, 63 (24 May 1833). — Type: “à l’Île Maurice ”, Bélanger s.n. (holo-, P00349575!).
Nephrodium filix-mas View in CoL auct. non (L.) Rich., Flora of Mauritius and the Seychelles: 496 (1877), pro parte.
OTHER MATERIAL EXAMINED. — Mauritius. Barkly & Lady Barkly s.n. (BM000801021, BM000801022).— On rocky side of Piton at Ripailles, Barkly & Lady Barkly s.n. (BM000801027). — Île Maurice, IX-X.1849, Boivin s.n. (P00349594, P00349595). — Île de France, Commerson s.n. (P00349589). — Mauritius, Corps de Gardes, Pike s.n. (NY). — Bois de Moka à l’Île de France, sine coll. s.n. (P00349590, 2 sheets).
Sine Loco. Barkly & Lady Barkly s.n. (BM000801028); Boivin s.n. (P00349591, P00349592, P00349593); Boivin s.n. (BM000801046); Bojer s.n. (K); Bojer s.n. (M0065885); Duncan s.n. (BM000801025); Mc Gregor s.n. (BM000801023, BM000801029); sine coll. s.n. (P00349571); sine coll. s.n. (P00349587); sine coll. s.n. (BM000801024).
DESCRIPTION
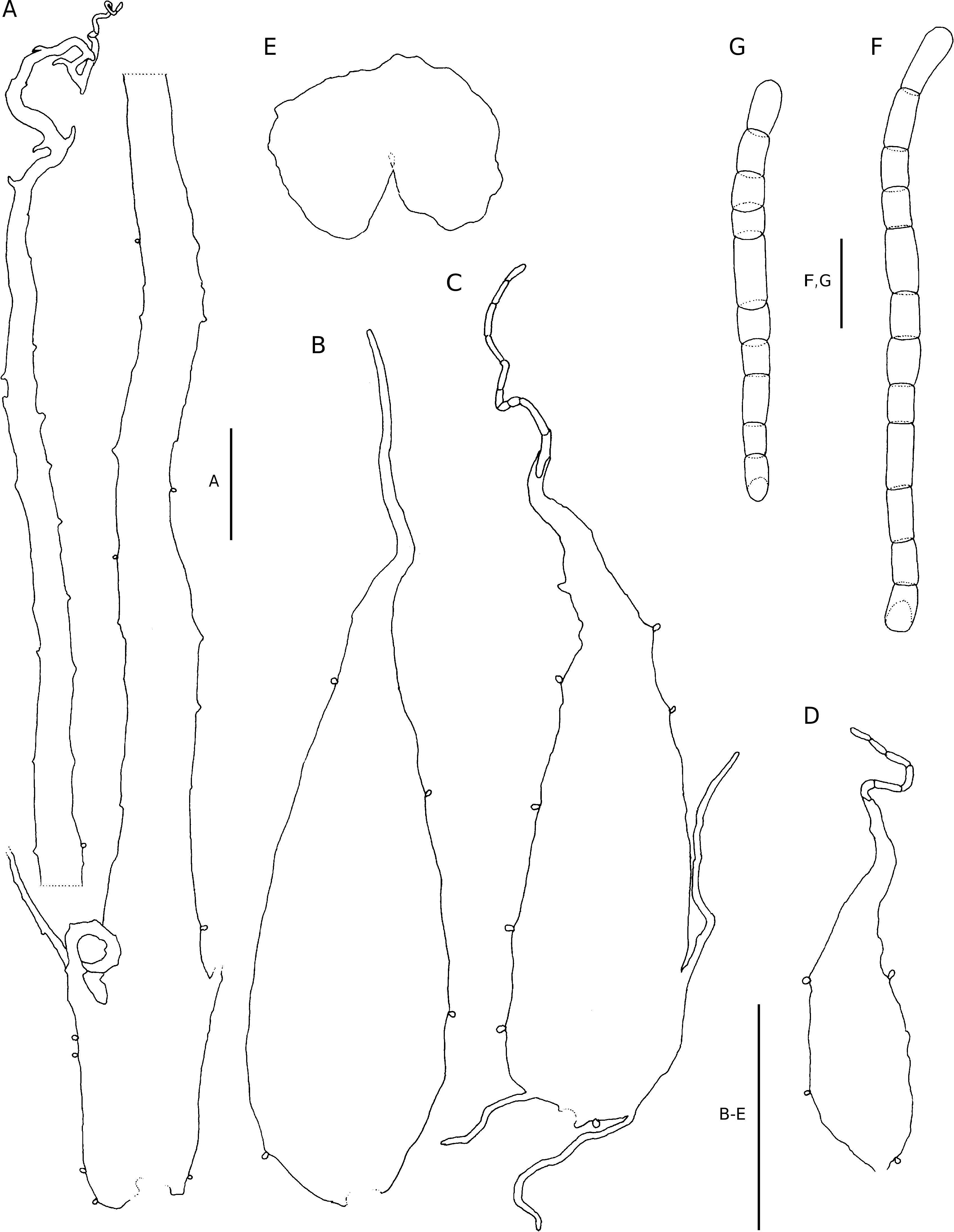
Plants terrestrial. Rhizome unknown. Fronds up to 1.2 m long; stipe stramineous, sulcate adaxially, up to c. 490 mm long and 6 mm in diameter, initially sparsely to moderately scaled, proximally densely scaled, subglabrous later, the scales ferrugineous, chartaceous, linear-attenuate, up to 32 × 5 mm, adnate, variously set with glands and long uniseriate pluricellular filiform outgrowths along the margin, the apex filiform; lamina anadromous, catadromous towards apex, broadly ovate to deltoid, up to 580 × 620 mm, to 3-pinnate, with up to 13 petiolated pinna pairs, distal pinnae become sessile and eventually adnate and increasingly basiscopically decurrent; rachis stramineous, adaxially sulcate, narrowly winged near the apex, sparsely scaled, the scales stramine- ous, thinly chartaceous, sessile, narrowly lanceolate to subulate, up to 4 × 1 mm, narrowly to broadly cuneate, often irregularly set with glands and long uniseriate pluricellular outgrowths, the apex terminates in a series of oblong cells; pinnae petiolate, the petiole up to 4 mm long, spaced or overlapping, near opposite to alternate, the basal pinna pair longest, to 2-pinnate, basal pinnae basiscopically developed, inaequilaterally lanceolate, up to 370 × 135 mm, pinnae towards the apex inaequilaterally oblong, with up to 10 petiolated pinnule pairs; pinna-rachis stramineous, adaxially sulcate, narrowly winged towards the apex, rarely sparsely glandular at the base, glands capitate, up to 62 µm long, sparsely scaled, the scales similar to those on the rachis; pinnules petiolate, the petiole up to 2 mm long, proximally spaced, slightly overlapping towards the pinna apex, the proximal basiscopic pinnule longest or not, acroscopic pinnule on basal pinnae ovate-acuminate to oblong-acuminate, up to 57 × 26 mm, pinnatifid, basiscopic pinnule on basal pinnae inaequilaterally lanceolate to oblong-acuminate, up to 105 × 34 mm, to 1-pinnate; pinnule-rachis stramineous, adaxially sulcate, pronounced abaxially, sparsely set with scales and hairs, the scales similar, but smaller than those on the pinna-rachis, the hairs isocytic; segments herbaceous, petiolate, the petiole up to 1 mm long, narrowly ovate to oblong-obtuse, up to 18 × 9 mm, glabrous adaxially, abaxially sparsely set with isocytic hairs along the costule and veins, hairs often long and long-celled, often glandular near the base, larger segments shallowly lobed, the lobes denticulate. Venation evident, lateral veins in pinnule lobes pinnately branched, vein branches end in the teeth near the margin. Stomata mostly of the polocytic type, (34-)45(-54) µm long. Sori essentially 2-seriate on larger pinnule lobes, discrete, supramedial on predominantly anadromous vein branches, up to 1 mm in diameter at maturity; indusium dark brown, firmly herbaceous, reniform, entire or shallowly repand, strongly recurved.Sporangium stalk simple, glandular, or haired, capsule with (14-)15(-16) indurated annulus cells, epistomium 4-celled, hypostomium 4-celled. Spores brown, 64 per sporangium, perispore folded to form pronounced reticulate ridges, exospore (32-)34(-44) × (18-)20(-22) µm.
DIAGNOSTIC FEATURES AND RELATIONSHIPS Diagnostic of the species are the scales, which bear numerous exceptionally long filiform outgrowths along their margins.
DISTRIBUTION AND HABITAT
Dryopteris bojeri is endemic to Mauritius and appear not to have been collected there recently ( Fig. 13 View FIG ). Lorence (1978) does not list the species, but he may have identified it as D. aquilinoides . Nothing is known about the ecology of the species.
4. Dryopteris comorensis (Tardieu) Fraser-Jenk. ( Figs 14 View FIG ; 15 View FIG )
Thaiszia 4 (1): 34 (1994) . — Dryopteris inaequalis (Schltdl.) Kuntze var. comorensis Tardieu, Notulae Systematicae (Paris) 15 (2): 162 (Apr. 1956). — Type: Comores, environs de Nioubadjou, Humblot 254 (holo-, BM000801045!; iso-, K!, P00220415!).
OTHER MATERIAL EXAMINED. — Comoro Islands. Grande Comore, à 5 km au Sud-est de Nioumbadjou, 600 à 700 m d’alt., 23.XI.1975, Coulon 277 (MO4374549). — Village de Kourani, 770 m, 11°51’S, 43°25’E, 15.II.1975, Floret 950 (MO4374543, P00220424). — Centre-Sud, entre Kourani et Dombéni vers 500 m d’alt., 15.II.1975, Floret 957 (P00220421). — Environs de Nioumbadjou , 400-500 m, IV.1911, Humblot s.n. (P00220417). — Comore Islands , 1884, Humblot 254 (BM000801045, K, P). — Massif du Kartala, versant ouest, 11°45’S, 43°19’E, 1900 m, forêt dense humide de montagne, Rakotondrainibe 6815 (P00312090, P00312091, P00312092) GoogleMaps .
Madagascar. Antsiranana, Parc national de la Montagne d’Ambre, environs du Lac Texier, rive NO. 1070 m, 27.VI.1992, Rakotondrainibe 1739 (P00100256) . — Antsiranana, Parc national de la Montagne d’Ambre, environs du lac Texier, rive NO, forêt primaire, cirque volcanique, 27.VI.1992, Rakotondrainibe 1740 (P00100257, P00100258, P00100259) .
DESCRIPTION
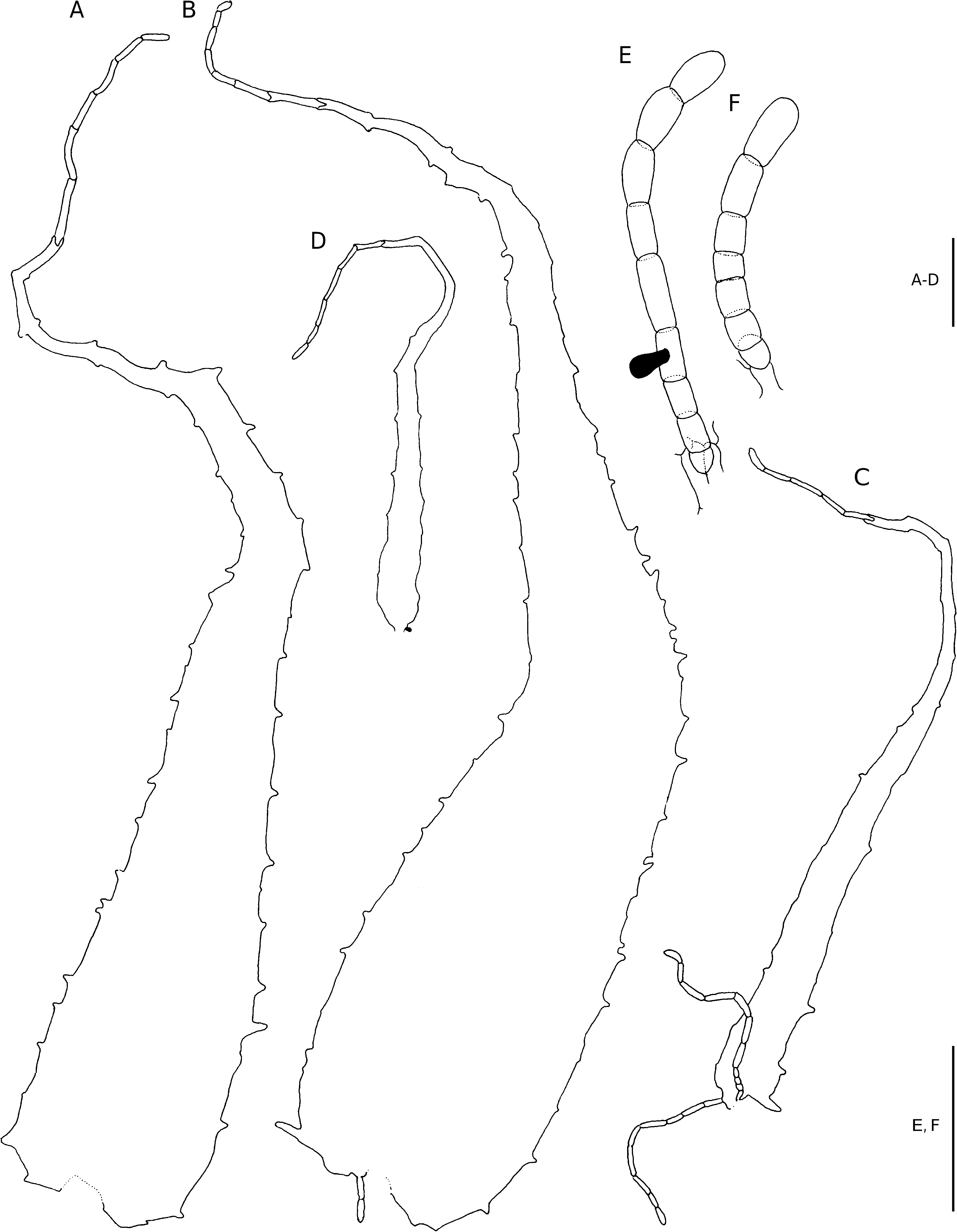
Plants terrestrial. Rhizome decumbent. Fronds crowded, suberect to arching, up to 1.9 m long; stipe proximally castaneous, stramineous higher up, sulcate adaxially, up to 830 mm long and 15 mm in diameter at the base, proximally densely scaled, moderately to sparsely scaled higher up, the scales ferrugineous, chartaceous, rugose, linear- acuminate, up to 30 × 5 mm, adnate, the margins regularly set with capitate glands and twisted pluricellular, mostly pluriseriate outgrowths, the apex filiform, twisted, terminating in an oblong cell; lamina anadromous, catadromous towards the apex, broadly ovate to deltoid, up to 1.1 × 0.78 m, to 3-pinnate, with up to 15 petiolated pinna pairs, pinnae near the apex become sessile and increasingly basiscopically decurrent; rachis stramineous, adaxially sulcate, initially moderately scaled, sparsely scaled later, the scales similar to, but smaller than those on the stipe, up to 12 × 3 mm; pinnae petiolate, the petiole up to 45 mm long, spaced or overlapping, near opposite to alternate, basal pinna pair longest, to 2-pinnate, basiscopically developed, inaequilaterally lanceolate, up to 440 × 225 mm, pinnae towards the apex oblong-acuminate, with up to 10 petiolated pinnule pairs; pinna-rachis stramineous, adaxially sulcate, narrowly winged towards the apex, sparsely to moderately scaled, the scales stramineous, thinly chartaceous, sessile or short-stalked, lanceolate-caudate to narrowly lanceolate-caudate, up to 13 × 2.3 mm, narrowly to broadly cuneate, irregularly set with glands along the margins and laminae, the margins also with long and twisted pluricellular outgrowths, the apex terminates in a series of oblong cells; pinnules petiolate, the petiole up to 2 mm long, proximally widely spaced or overlapping, proximal basiscopic pinnule longest or not, acroscopic pinnule on basal pinnae oblong-acuminate, up to 87 × 27 mm, 1-pinnate, basiscopic pinnule on basal pinnae oblong-acuminate, up to 135 × 45 mm, to 1-pinnate, with up to 6 petiolated segment pairs; pinnule-rachis stramineous, adaxially sulcate, pronounced abaxially, sparsely haired adaxially, sparsely to moderately scaled abaxially, the scales similar to those on the pinna-rachis, up to 5 × 1.4 mm; segments herbaceous, petiolate, the petiole up to 0.5 mm long, sessile and increasingly basiscopically decurrent along the costa towards the apex, narrowly lanceolate to oblong-acuminate, up to 27 × 10 mm, lobed, the lobes oblong-obtuse, up to 5 × 4 mm, shallowly denticulate, adaxially with a few hairs along the costule, abaxially sparsely to moderately set with hairs and scales along the costule and veins, the hairs isocytic, up to 2.5 mm long, the scales stramineous to ferrugineous, thinly chartaceous, stalked, subulate-caudate, up to 2.5 × 0.6 mm, the apex terminates in a series of oblong cells. Venation evident, lateral veins in segment lobes pinnately branched, vein branches end in the teeth near the margin. Stomata mostly of the polocytic type, (34-)40(-46) µm long. Sori essentially 2-seriate on segments, discrete or touching at maturity, medial on predominantly anadromous vein branches, up to 1 mm in diameter at maturity; indusium ferrugineous, firmly herbaceous, reniform, entire or shallowly repand, strongly recurved. Sporangium stalk simple, glandular, or haired, capsule with (13-)15(-18) indurated annulus cells, epistomium (3-)4(-6)-celled, hypostomium 4(-6)-celled. Spores brown, 64 per sporangium, exospore (34-)37(- 44) × (20-)23(-26) µm.
DIAGNOSTIC FEATURES AND RELATIONSHIPS Dryopteris comorensis is separated from other species in the Mascarene region by the rugose scales, which form a dense covering on the lower half of the stipe. These scales bear capitate glands and numerous long and twisted outgrowths along the margins. Although similar marginal outgrowths may occur in D. pentheri , that species does not occur on Grande Comore, neither does it bear unicellular oblong glands or 2-celled hairs along the lamina axes and veins. Scales along the pinnarachis are often somewhat bullate. The stoma size is also smaller, (34-)40(-46) µm, which suggests it being diploid, rather than (34-)53(-72) µm as in D. pentheri , which is tetraploid.
Fraser-Jenkins (1986) failed to include the taxon in his classification. I see no evidence of this species being related to Dryopteris inaequalis (Schltdl.) Kuntze , as suggested by viewing it as a variety of this species (Tardieu-Blot 1956a: 162). I support Fraser-Jenkins (1994) in recognizing it as a distinct species.
DISTRIBUTION AND HABITAT
Dryopteris comorensis occurs on Grande Comore, also known as Ngazidja (11°45’S, 43°19’E), and on Montagne d’Ambre (c. 12°32’S, 49°10’E), in northern Madagascar ( Fig. 16 View FIG ). The species is restricted to the evergreen rain forests and occurs at altitudes ranging between 400 to 1900 m on Mount Kartala. Both Mount Kartala and Montagne d’Ambre are of volcanic origin GoogleMaps .
5. Dryopteris manniana (Hook.) C.Chr. ( Figs 17 View FIG ; 18 View FIG )
Index filicum : 276 (18 Nov. 1905). — Polypodium mannianum Hook. , Species filicum 4 (15/16): 253 (1863). — Phegopteris manniana (Hook.) Kuhn, Filices africanae: 123 (Oct. 1868). — Type: Fernando Po, on the peak, 2000 ft, 1860, G. Mann s.n. (holo-, K!; iso-, K!).
OTHER MATERIAL EXAMINED. — Madagascar. Ambatofitorahana, 1700 m, IX.1956, Bosser 9787 (P00349514).— Centre, Forêt de Manjakatompo , versant E de l’Ankaratra, VII.1953, Capuron 70RC (P00349509). — Rahobevava, 960 m, 11.III.1951, Cours 4292 (P00349513).— Madagascar, Cowan s.n. (BM000800961, A only, BM000800962), Ankafana, Cowan s.n. (BM000605571). — Madagascar, Cowan s.n. (BM000605576). — Prov. Imerina, VII.1880, Hildebrandt 3529 (BM000800984). — Massif du Kalambatitra (Centre-Sud), Mont Kalambatitra et ses abords et mont Analatsitendrika, forêt ombrophile (sur latérite de gneiss), 1500-1750 m, XI.1933, Humbert 11875 (P00349512). — Tanala, V.1880, Kitching s.n. (K, B only). — Antananarivo, Ankazobe, Manankazo, réserve spéciale d’Ambohitantely, environ 8 km au Nord- Ouest d’Ankazobe, 18°10’S, 47°17’E, 1200-1650 m, Rakotondrainibe 348 (P00349515) GoogleMaps .
DESCRIPTION
Plants terrestrial.Rhizome erect to short-decumbent, mostly unbranched, up to 8 mm in diameter, set with roots, closely spaced stipe bases, and scales, the scales brown to ferrugineous, chartaceous, broadly attached, subulate, up to 22 × 3 mm, the margins closely set with short teeth, the apex filiform, terminating in an oblong cell. Fronds 4-7 per plant, caespitose, arching, up to 1.0 m long; stipe greenish to stramineous, shallowly sulcate adaxially, up to 480 mm long and 5 mm in diameter, proximally densely scaled, moderately scaled higher up, the scales stramineous to ferrugineous, chartaceous, the larger scales broadly attached, up to 15 × 4.5 mm, the smaller scales short-stalked, cordate to cuneate, the margins denticulate, basally frequently with one or more filiform outgrowths, often with scattered glands, the apex filiform, terminating in an oblong cell; lamina herbaceous, ovate, up to 525 mm long, to 2-pinnate-pinnatifid, anadromous, catadromous towards the apex, with up to 12 petiolated pinna pairs, proliferous, generally with one or more scaled buds adaxially along the rachis near the lamina apex, often also with buds on the pinna-rachis in larger fronds; rachis greenish to stramineous, adaxially sulcate, narrowly winged towards the apex, moderately scaled, the scales ferrugineous to stramineous, chartaceous, sessile or short-stalked, filiform to lanceolate, up to 7 × 1.8 mm, cordate to narrowly cuneate, denticulate, basally frequently with one or more filiform outgrowths, higher up closely set with short teeth, the apex filiform, terminating in an oblong cell; pinnae petiolate, the petiole up to 18 mm long; basal pinna pair inaequilaterally ovate, narrowly ovate to oblong-acuminate towards the lamina apex, up to 250 × 95 mm, to 1-pinnatepinnatifid; basal pinna pair mostly the longest, basiscopically developed, opposite to alternate, basally widely spaced, more closely spaced towards the lamina apex and often imbricate, with up to 3 pairs of petiolated pinnules; pinna-rachis adaxially shallowly sulcate, the sulcus confluent with that of the rachis, pronounced abaxially, narrowly winged for most of the length, abaxially sparsely to moderately scaled, the scales stramineous to ferrugineous, chartaceous, sessile or short-stalked, filiform to narrowly oblong, up to 5 × 1 mm, cordate to narrowly cuneate, dentate, basally frequently with one or more filiform outgrowths, the apex filiform, terminating in an oblong cell; pinnules petiolate, the petiole up to 2 mm long, symmetric or inaequilaterally narrowly ovate to ovate, up to 50 × 17 mm, basiscopically decurrent, pinnatifid, acroscopic pinnule on the basal pinnae up to 56 × 23 mm, not or slightly imbricate, the lobes broadly oblong-obtuse, shallowly lobed, dentate, glabrous adaxially, abaxially rarely glandular, the glands (56-)62(-72) µm long, sparsely set with hairs and scales along the veins, the hairs moniliform, often with a unicellular gland near the base, the scales stramineous, chartaceous, short-stalked, the stalk often glandular, filiform to linear, up to 2 mm long, repand to dentate, the apex filiform, terminating in an oblong cell. Venation anadromous, becoming catadromous towards the lamina and pinna apex, pinnately branched in the lobes, evident, ending in the teeth near the margin, the endings mostly slightly enlarged. Stomata mostly of the polocytic type, (40-)56(-70) mm long. Sori circular, inframedial on unmodified fertile vein branches, discrete at maturity, up to 1.5 mm in diameter; sporangium stalk simple or haired, capsule with (11-)14(-18) indurated annulus cells; exindusiate. Spores ellipsoidal, monolete, perispore forming short and long ridges, ruminate, (40-)44(- 58) × (26-)30(-32) mm. Chromosome number: 2n = c. 164 (Vida in Widén et al. 1973).
DIAGNOSTIC FEATURES AND RELATIONSHIPS Diagnostic of Dryopteris manniana is the scaled proliferous buds borne adaxially on the rachis, mostly near the lamina apex, the denticulate scales, and the exindusiate sori. A micromorphological feature separating it from other Dryopteris species is the stoma size, which supports it being tetraploid.
VARIATION
Morphologically, the species appears to be stable throughout its distribution, with little variation of significance having been observed.
DISTRIBUTION AND HABITAT
Dryopteris manniana is widespread in sub-Sahara Africa occurring in West-, East-, southern Africa, and in Madagascar. It chiefly occurs in moist montane forests, at elevations ranging from 1400 m in Kenya and Uganda to 2249 m along the Kikuyu Escarpment in Kenya. In Madagascar, it is known from elevations ranging between 960 and 1750 m ( Fig. 16 View FIG ).
6. Dryopteris pentheri (Krasser) C.Chr. ( Figs 19 View FIG ; 20 View FIG )
Index filicum : 284 (18 Nov. 1905). — Nephrodium pentheri Krasser, Annales des Kaiserlich-Königlichen Naturhistorischen Hofmuseums , Wien 15: 5, 6 (1900). — Type: South Africa, Van Reenens Pass , 4.III.1895, F. Krook s.n., sub Penther Plantae Austro-Africanae 36 (lecto-, W8042! designated by Pichi Sermolli [1984: 329]; isolecto-, BM!),.
Dryopteris inaequalis sensu Tardieu, Flore de Madagascar et des Comores (Plantes Vasculaires), 5e Famille. Polypodiacées (sensu lato) 5 (1) Dennstaedtiacées. (10) Aspidiacées ): 315 (May 1958), pro parte, non (Schltdl.) Kuntze (1891: 813).
MATERIAL EXAMINED. — Madagascar. Route d’Anjozorobe, prés d’Analabe, XII.1958, Bosser 12329 (MO4405417). — Route d’Arivonimamo, 1958, Bosser 12344 (MO4405414). — Andramasina, XII.1958, Bosser 12446 (MO4405413).— Route de Manankaraly, III.1957, Bosser 12871 (MO4405415). — Ankaratra, V.1962, Bosser 16183 (MO4405425). — P.K. 39 route du sud, ravin humide, III.1953, Bosser 6203 (MO4405416). — Imarina, 1881, Cowan s.n. (BM000800959, BM000800960, BM000800981, BM000800985, BM000800998, BM000801020, BM000801031, BM000801044). — Fianarantsoa, 1879, Cowan s.n. (BM000801030).— Prov. Tananarive, vicinity of Station forestière Antsampandrano, disturbed prairie, 1750 m, 23.I.1975, Croat 29124 (MO3295933). — Prov. Fianarantoa, forest and forest edge along Route 7, 113- 116 km N of Fianarantsoa, 1580-1590 m, 28.I.1975, Croat 29928 (MO3295935). — Massif de l’Anakaratra, flanc oriental duTsiafajavona, reste de forêt, 1700-2000 m, 15.VII.1928, Decary, Humbert & Swingle 4568 (BM000801032). — Interior, near waterfall Ramainandro, Elliot 1965 (K). — Ambohimitombo forest, 19.I.1895, Forsyth Major 180 (BM000800986, K). — Prov. Imerina, IV.1884, Hildebrandt 3529 (K, M, US815398). — Madagascar, V.1880, Kitching s.n. (K, A only). — Ilafy, Tananarive, 6.I.1970, Onreadt 70M66 (US3005426). — Moramanga, c. 900 m, Regenwald, Schlieben 8109 (M). — La Mandraka, c. 1000 m, Regenwald, 7.XII.1959, Schlieben 8137 (BM, K, M, 2 sheets, PRE, US3001350). — Bord de l’eau à l’ombre Tsimbazaza, 14.VII.1885, sine coll. s.n. (P00349496). — Madagascar, sine coll. 68 (NY, US719837). — Madagascar, sine coll. 87 (NY, US719841). — Province du Vakinankaratra, Distr. d’Antsirabe, fentes des rochers sur la pente interne du cratère du Tritriva vers 1800 m d’alt., 16.XI.1972, Viguier & Humbert 1294 (P00349498).
Réunion. Fissures aux environs du Piton Maido, Grand Bénard, 2200 m, 22.XI.1968, Cadet 1751 (P00349598). — Sous-bois de la forêt hygrophile, forêt de la Mare à Joseph Cilaos, 1400 m, 7.II.1969, Cadet 1904 (P00349599, P00349600).— Clairière dans la forêt du Grand Matarum Cilaos, 1600 m, 23.III.1974, Cadet 4582 (P00349596, P00349597). — Cilaos Distr., footpath from Cilaos thermal baths to Bras Rouge cataract, on earth bank with Polystichum ammifolium, 1100 m , 4.X.1984, Jacobsen 5608 (FR).
DESCRIPTION
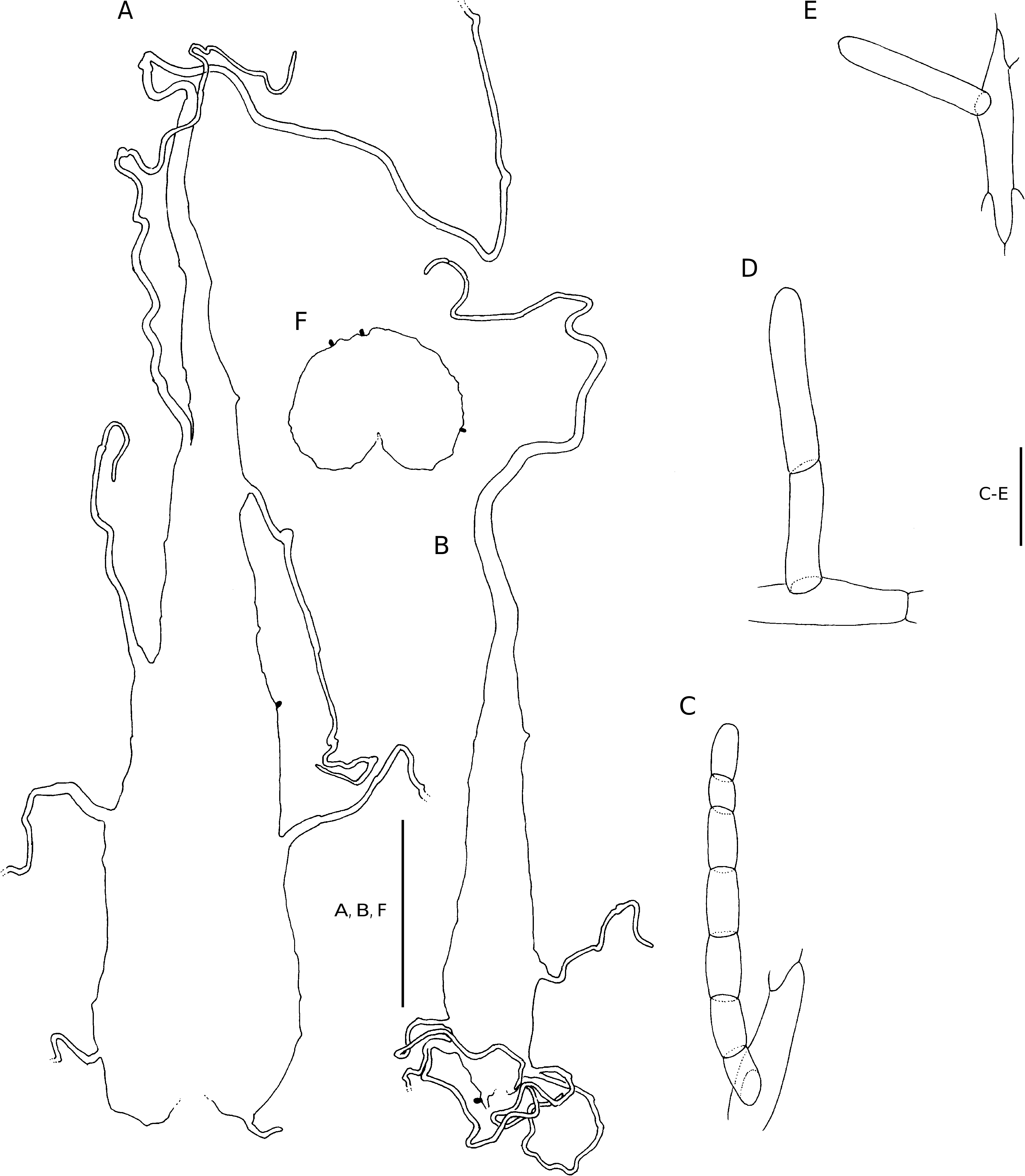
Plants terrestrial or epilithic. Rhizome short-decumbent, sparsely branched, up to 20 mm in diameter, set with roots, closely spaced stipe bases, and scales, the scales stramineous to ferrugineous, chartaceous, broadly attached, linear, oblong or narrowly ovate, up to 37 × 6 mm, truncate, the margins irregularly set with long, twisted, pluricellular filiform outgrowths, the apex filiform, twisted. Fronds caespitose, erect to arching, up to 1.8 m long; stipe proximally castaneous, brown to stramineous higher up, proximally adaxially flattened, shallowly sulcate higher up, up to 485 mm long and 10 mm in diameter, proximally densely scaled, the scales higher up fugaceous, stramineous to ferrugineous, chartaceous, the larger scales up to 40 × 7 mm, broadly attached, the smaller scales sessile or short-stalked, narrowly lanceolate to filiform, cordate to cuneate, the margins irregularly set with long, twisted, pluricellular filiform outgrowths and often also with scattered glands, the apex filiform; lamina herbaceous to firmly herbaceous, ovate to ovate-triangular, up to 780 mm long, 2-pinnate to 3-pinnate, anadromous, catadromous towards the apex, with up to 16 petiolated pinna pairs; rachis stramineous, adaxially shallowly sulcate, becoming narrowly winged towards the apex, initially moderately scaled, the scales fugaceous, broadly attached or short-stalked, narrowly lanceolate to linear, up to 7 × 2 mm, cuneate, the margins irregularly set with long filiform outgrowths, often also with scattered glands, the apex filiform, twisted; pinnae petiolate, the petiole up to 18 mm (rarely to 38 mm) long, the basal pinna pair inaequilaterally ovate to narrowly ovate, narrowly lanceolate to oblong-acuminate towards the lamina apex, to 2-pinnate, basal pair the longest, mostly basiscopically developed, up to 315 × 185 mm, opposite to alternate, basally widely spaced, often somewhat imbricate higher up, with up to 9 petiolated pinnule pairs; pinna-rachis adaxially shallowly sulcate, the sulcus confluent with that of the rachis, pronounced abaxially, narrowly winged distally, abaxially moderately to sparsely set with scales and hairs, the scales and hairs stramineous to ferrugineous, the scales chartaceous to membranous, broadly attached or short-stalked, narrowly to broadly cuneate, up to 5 × 1.6 mm, the margins irregularly set with long pluricellular, filiform outgrowths, the apex filiform, twisted, up to 5 × 1.6 mm, the hairs are of three types: 1) unicellular oblong glands, 2) bicellular hairs with straight or oblique transverse walls, and 3) pluricellular simple or branched, mostly isocytic hairs often bearing a single glandular cell near the base; pinnules petiolate, petiole up to 3 mm long, aequilaterally to inaequilaterally lanceolate to oblongacuminate, basiscopically decurrent, 1-pinnate to lobed, acroscopic pinnule on the basal pinnae up to 90 × 35 mm, basiscopic pinnule on the basal pinnae up to 112 × 42 mm, widely spaced or imbricate; pinnule-rachis adaxially shallowly sulcate, pronounced abaxially, narrowly winged, the wing continuous with that of the pinna-rachis, variously set with scales, isocytic- and 2-celled hairs; segments widely spaced to imbricate, ovate to oblong-obtuse, up to 22 × 9 mm, basiscopically decurrent, lobed, the lobes serrate, adaxially glabrous, with oblong glands along and between the veins, or with a few pluricellular mostly isocytic hairs along the costule, abaxially sparsely set with unicellular oblong glands (60-)137(-260) mm long, 2-celled hairs, and isocytic and/or moniliform hairs, those near the segment base often with a glandular cell near the base, and filiform scales, the scales short-stalked, the stalk often with one or more glandular cells, hairs and scales mostly occur along the costule and veins. Venation anadromous, becoming catadromous towards the lamina and pinna apex, pinnately branched in the segments, vein branches pinnately branched, forked or simple near the apex, evident, ending in the teeth near the margin, endings slightly enlarged and often conspicuous adaxially. Stomata of the anomo- and polocytic types, (34-)53(-72) mm long. Sori circular, medial on predominantly anadromous vein branches, discrete, up to 1.8 mm in diameter at maturity, essentially uniseriate; sporangium stalk simple, or with one or more glandular cells, but mostly with a long multicellular, uniseriate hair, capsule with (11-)13(-20) indurated annulus cells, epistomium (3-)6(-8)-celled, hypostomium (3-)6(-8)-celled; indusium persistent, pale brown, firmly herbaceous, reniform, entire, repand, or erose, (rarely glandular along margin), often strongly revolute, to 1.8 mm in diameter. Spores ellipsoidal, monolete, perispore folded into tubercules or reticulate ridges, finely rugose to ruminate, (38-)45(-60) × (27-)31(-40) mm (Tryon & Lugardon 1990: 426, fig. 159.21). Chromosome number: 2n = c. 164 (Vida in Widén et al. 1973: 2129).
REMARKS
The spores of Viguier & Humbert 1731 (P00349497) are highly abnormal and may be indicative of it being a hybrid. Other features of the specimen – stoma size and the presence of 2-celled hairs – are typical of D. pentheri .
DIAGNOSTIC FEATURES AND RELATIONSHIPS Dryopteris pentheri is an exteremely variable species difficult to separate from other species using
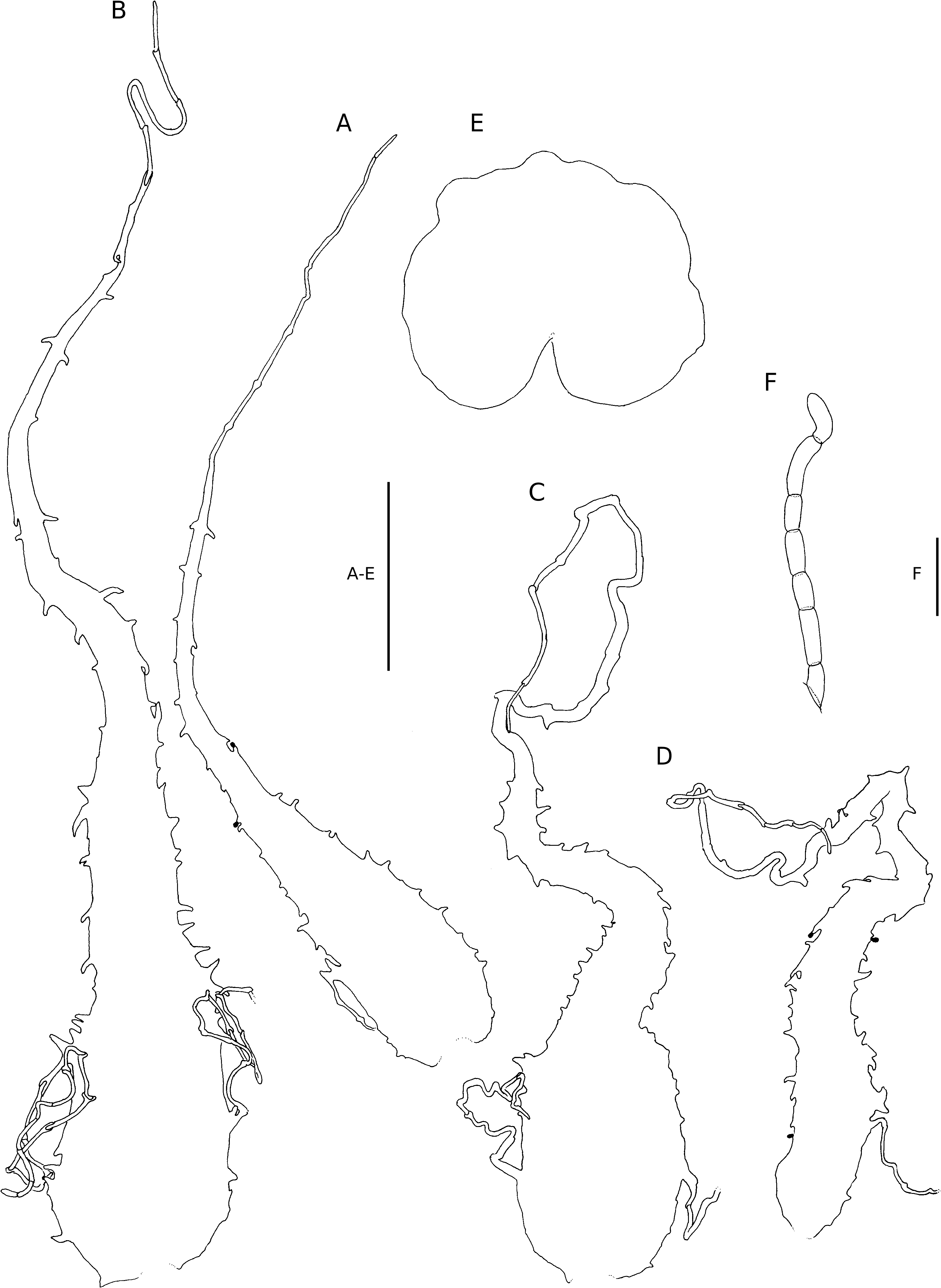
macromorphological features. Micromorphological features separating it from other Dryopteris species in the region are the oblong glands ( Fig. 19E View FIG ) and 2-celled hairs ( Fig. 19D View FIG ), occurring along the frond axes and veins, and the larger stomata ( Table 1).
VARIATION
Dryopteris pentheri is extremely variable in gross morphology and attempts by several authors to separate this species based on macromorphological characters have failed. Micromorphological characters and cytological observations are more reliable in separating D. pentheri from other taxa. Micromorphological features of significance are the presence of bicellular hairs, unicellular gland length, as well as stoma and spore size. These features, however, all show some variation. Bicellular hairs occurring abaxially on the lamina were observed on the costa, costules or veins in 64.5% of the collections studied. These hairs mostly occur centrally on an epidermal cell, but rarely also near the distal end of a cell. The periclinal wall of the epidermal cells bearing the hairs is flat or the hairs are positioned on a low but prominent bulge. The transverse wall between the two hair cells may be straight or oblique. Unicellular glandular hairs were observed in 96.8% of the collections studied. These oblong hairs, 60-260 mm long, are positioned on the epidermal cells in a similar fashion as bicellular hairs. Pluricellular hairs 6 to 21 cells long occur adaxially and abaxially on or near the costae, costules and veins. They are of the isocytic or moniliform type and are positioned centrally on or near the distal margin of an epidermal cell. They often bear a single (rarely 2) glandular cell near the base. The indusium also shows significant variation in size and margin sculpture. In a few cases, glands have been observed along the indusium margin.
DISTRIBUTION AND HABITAT
Dryopteris pentheri is widespread in the eastern and western mountainous regions of sub-Sahara Africa, Madagascar and Réunion ( Fig. 20 View FIG ). Although predominantly confined to moist forests, the species occurs in open habitats at higher elevations. In these habitats, the plants are mostly confined to boulder bases and rock crevices on ridges and montane streams. In grassland habitats, the species is exposed to regular burning, but this appears to have little effect on the subterranean rhizome. In Madagascar, it is confined to the central mountainous regions, at elevations ranging between 1580 and 2000 m, but on Réunion, it occurs in moist montane forests and in seasonally wet forests from 1100 to 2200 m, where the plants often become seasonally dormant.
7. Dryopteris subcrenulata (Baker) C.Chr. ( Figs 22 View FIG ; 23 View FIG )
Index filicum : 295 (18 Nov. 1905). — Nephrodium subcrenulatum Baker View in CoL , Journal of the Linnean Society, Botany 16: 202 (1877). — Aspidium subcrenulatum (Baker) Kuhn, Von der Decken’s Reisen in Ost-Afrika in 1851-1861 3 (3): 65 (Aug.-Sep. 1879). — Type: Madagascar, Antananarivo, H. Gilpin s.n. (holo-, K000351090!, K000351091!).
Dryopteris mangindranensis Tardieu, Notulae Systematicae (Paris) 15 (2): 161, 162 (Apr. 1956). — Type: Madagascar, montagnes au Nord de Mangindrano , jusqu’aux sommets d’Ambohimirahavavy, 1900-2500 m, Humbert 25054 (holo-, P00483232!; iso-, K000351050!,).
OTHER MATERIAL EXAMINED. — Madagascar. Ambohimitombo forest, 27.XI.1894, Forsyth Major 193 (K). — Montagnes au Nord du Mangindrano (haute Maevarano), jusqu’au sommet d’Ambohimirahavavy (partage des eaux Mahavavy-Androranga: Centre-Nord): haute Bemafo (affluent de l’Androranga), forêt ombrophile sur latérite de gneiss, 1900 m, Humbert & Capuron 25054 (P00349516). — Montagnes au Nord de Mangindrano (haute Maevarano) jusqu’aux sommets d’Ambohimirahavavy (partage des eaux Mahavavy- Androranga: Centre-Nord), 1800-2000 m, 19.I-12. II.1951, Humbert & Capuron 25055 (P00349518). — Massif de Marivorahona au sud-ouest de Manambato (haute Mahavavy du nord, district d’Ambilobe), forêt ombrophile sur gneiss, 1750-2000 m, 18-26.III.1951, Humbert & Capuron 25810 (P00349519).— Tampoketsa au N d’Ankazobe (Centre), forêt d’Ambohitantely et reste de forêts aux alentours, vers 1600 m, 1933, Humbert 11127 (P00349502). — Massif du Kalambatitra (Centre-Sud), Mont Beanjavidy: forêt ombrophile, 1500- 1730 m, XI.1933, Humbert 12064 (P00349520). — Mt Tsaratanana, 2000 m, Humbert 16165 (P00349510). — Bassin supérieur du Sambirano, forêt ombrophile sur sol siliceux, 1700 m, XI-XII.1937, Humbert 18656 (P00349511). — Mt Tsaratanana, 2000 m, IV.1923, Perrier de la Bathie 16458 (P00349517). — Massif de Manongarivo, bois vers 800 m, III.1909, Perrier de la Bathie 7445 (P00349501). — Antananarivo, 4/76, Pool s.n. (K). — Antsiranana, Andapa, RNI 12 du Marojejy, au bord d’un affluent de la rivière Manantenina, à 10 km au Nord-Ouest du village de Manantenina, 820 m, 20.X.1996, Rakotondrainibe 3429 (P00084909, P00084910). — Toamasina, near Andisabe, forest of Mantadia, beyond the graphite mine, 900 m, 3.XI.1994, Van der Werff, McPherson & Rapanarivo 13626 (PRE).
DESCRIPTION
Plants terrestrial. Rhizome short-decumbent to suberect, up to 110 mm long and 15 mm in diameter, set with roots, closely spaced stipe bases and scales, the scales ferrugineous, thinly chartaceous, broadly attached, lanceolate to subulate, up to 15 × 3.5 mm, proximally entire, denticulate towards the apex, often with scattered glands and/ or one or more pluricellular, mostly recurved outgrowths along the margin, the smaller scales often with more marginal outgrowths, the apex filiform, terminating in a uniseriate series of oblong cells. Fronds caespitose, suberect to arching, up to 7 per plant, up to 1.2 m long; stipe up to 640 mm long and 8 mm in diameter, proximally castaneous, stramineous higher up, sulcate adaxially, initially densely scaled, the scales fugaceous, castaneous, chartaceous, the larger scales broadly attached, ovate to lanceolate, up to 15 × 3.4 mm, cordate to broadly cuneate, denticulate, often irregularly glandular along the margins, and/or with one or more mostly recurved pluricellular outgrowths, the apex filiform, terminating in a series of oblong cells, the smaller scales short-stalked, the stalk often glandular, subulate, narrowly cuneate, denticulate, the apex terminating in a short series of oblong cells; lamina ovate to broadly ovate, 1-pinnatepinnatifid to 2-pinnate, up to 710 × 420 mm, with up to 16 petiolated pinna pairs, the basal pinnae widely spaced, becoming increasingly more closely spaced distally, never overlapping; rachis stramineous, sulcate adaxially, narrowly winged towards the apex, sparsely scaled, the scales stramineous to ferrugineous, chartaceous, short-stalked, linear to subulate, up to 11 × 2 mm, entire, irregularly denticulate, often with scattered glands along the margins and with numerous filiform outgrowths, the apex terminates in a short series of oblong cells; pinnae petiolate, the petiole up to 4 mm long, near opposite to alternate, becoming increasingly basiscopically decurrent along the rachis and more broadly attached towards the lamina apex, falcate, herbaceous, one to two basal pinna pairs inaequilaterally oblong-acuminate to narrowly lanceolate, pinnatifid to 1-pinnate, symmetrically oblong-acuminate towards the lamina apex, becoming less deeply lobed towards lamina and pinna apices, basal pinnae longest or not, up to 260 × 62 mm, with up to 8 petiolated pinnule pairs; pinna-rachis shallowly sulcate adaxially, glabrous, pronounced abaxially, narrowly winged for most of the length, sparsely scaled, the smaller scales stramineous to ferrugineous, chartaceous, shortstalked, the stalk often glandular, filiform, up to 5 × 1.4 mm, denticulate or with several filiform marginal outgrowths, the apex terminates in a short series of oblong cells, the larger scales sessile, cordate to cordate-imbricate; pinnules sessile or petiole up to 1 mm long, acroscopic pinnule on basal pinnae oblong-obtuse to oblong acute, up to 28 × 13 mm, basiscopic pinnule on basal pinnae up to 24 × 10 mm, shallowly to deeply lobed, the lobes entire to shallowly crenulated to obtusely denticulate, oblong-obtuse, mostly widening acroscopically and basiscopically towards the lamina apex, up to 8 × 5 mm, glabrous adaxially, or with a few isocytic hairs near the costa, abaxially with isocytic hairs up to 0.8 mm long along and near the veins, hairs often glandular near the base. Venation evident, lateral veins in pinnules pinnately branched, forked or simple, vein branches end near the margin. Stomata mostly of the polocytic type, (34-)50(-58) mm long. Sori 2-seriate on lobes, medial to supramedial on predominantly anadromous vein branches, up to 1.4 mm in diameter; sporangium stalk simple or haired, capsule with 14(-15) indurated annulus cells, epistomium 4(- 5)-celled, hypostomium 4(-5)-celled. Indusium castaneous, firmly chartaceous, reniform, entire, up to 1.5 mm in diameter, margins strongly recurved, entire. Spores 64 per sporangium, brown, with short reticulate ridges and bulges, (32-)38(-42) × (22-)23(-24) mm.
DIAGNOSTIC FEATURES AND RELATIONSHIPS Characteristic of the species is the narrow rhizome and frond scales, which are denticulate and which often bear a few very long filiform outgrowths along the margins. The outgrowths are mostly uniseriate and are composed of several long cells. The joints between these cells are sinuate.
DISTRIBUTION AND HABITAT
A species confined to Madagascar, where it occurs in moist evergreen forests, at elevations ranging between 800 and 2000 m ( Fig. 24 View FIG ).











No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dryopteris bernieri Tardieu
| Roux, Jacobus P. 2011 |
Nephrodium filix-mas
| Rich. 1877: 496 |