
Metopograpsus cannicci, Innocenti & Schubart & Fratini, 2020
|
publication ID |
https://doi.org/ 10.26107/RBZ-2020-0077 |
|
publication LSID |
lsid:zoobank.org:pub:D535DC5B-050E-4982-924B-B8DF9C45A03F |
|
persistent identifier |
https://treatment.plazi.org/id/C4922D1A-6F0D-45EA-9AC9-659BDB005A1A |
|
taxon LSID |
lsid:zoobank.org:act:C4922D1A-6F0D-45EA-9AC9-659BDB005A1A |
|
treatment provided by |
Diego |
|
scientific name |
Metopograpsus cannicci |
| status |
sp. nov. |
Metopograpsus cannicci View in CoL , new species
( Figs. 1A View Fig , 2A View Fig , 3A View Fig , 4A View Fig , 5A View Fig , 6 View Fig )
Material examined. Types. Holotype. 1 male (34.5 × 28.1 mm) ( MZUF 4865 View Materials ), Egypt: Ras Mohammad, mangrove channel, colls. S. Fratini, G. Innocenti, S. Barbaresi & A. Conti, 12 October 2004 . Paratypes. 1 male (33.6 × 27.4 mm) ( MZUF 5010 View Materials ), Egypt: Nabq , El Arwashie, colls. S. Fratini, G. Innocenti, S. Barbaresi & A. Conti, 15 October 2004 ; 1 male (27.6 × 21.5 mm) ( ZRC 2019.1350 View Materials ), Nabq , El Monqata, colls. S. Fratini, G. Innocenti, S. Barbaresi & A. Conti, 13 October 2004 ; 1 male (24.4 × 19.2 mm) ( MZUF 5007 View Materials ), same collection data as ZRC 2019.1350 ; 1 female (24.4 × 19.4 mm) ( ZRC 2019.1351 View Materials ), same collection data as ZRC 2019.1350 ; 1 female (18.8 × 14.3 mm) ( MZUF 5011 View Materials ), same collection data as ZRC 2019.1350 .
Others. Egypt: 1 male (24.6 × 19.4 mm) , 1 female (21.8 × 16.6 mm) ( MZUF 5012 View Materials ), Egypt: Nabq, El Arwashie , colls. S. Fratini, G. Innocenti, S. Barbaresi & A. Conti, 15 October 2004 ; 1 male (18.9 × 14.0 mm) ( MZUF 5013 View Materials ), Ras Mohammad, Mangrove channel, colls. S. Fratini, G. Innocenti, S. Barbaresi & A. Conti, 14 October 2004 . Somalia: 1 male (23.3 × 17.8 mm) , 4 females (27.4 × 20.8 mm, 27.9 × 22.0 mm, 27.8 × 21.0 mm, 23.0 × 17.9 mm) ( MZUF 4935 View Materials ), Gesira, mangroves, coll. M. Vannini, 14 October 1981 ; 3 males (25.9 × 20.1 mm, 22.2 × 16.9 mm, 21.1 × 16.5 mm) ( MZUF 647 View Materials ), Gesira, coll. M. Vannini, December 1976 . Kenya: 2 males (24.9 × 19.0 mm, 25.0 × 19.1 mm) , 1 female (25.4 × 19.1 mm) ( MZUF 4940 View Materials ), Mida Creek , Dabaso, coll. S. Cannicci , December 1997 ; 1 male (22.1 × 17.8 mm), 1 female (25.5 × 19.6 mm) ( MZUF 4943 View Materials ), Mida Creek , Sita, coll. M. Vannini, October 1988 ; 1 female (27.1 × 21.0 mm) ( MZUF 4939 View Materials ), Mida Creek , coll. M. Vannini, October 1990 ; 1 female (25.2 × 19.2 mm) ( MZUF 3654 View Materials ), Mida Creek , Sita, coll. M. Vannini, July 2000 ; 1 male (25.8 × 20.3 mm) ( MZUF 4945 View Materials ), Mida Creek , Dabaso, coll. M. Vannini, 15 October 1990 ; 1 male (22.0 × 16.9 mm) ( MZUF 2544 View Materials ), Mida Creek , coll. V. Vigiani, September 2000 . Tanzania: 2 males (20.8 × 15.5 mm, 18.7 × 13.9 mm) ( MZUF 5003 View Materials ), Pangane , Mucojo, Macomia, mangrove swamp, coll. F. Tarducci, 26 August 1995 . Mozambique: 2 males (17.3 × 13.3 mm, 15.4 × 12.2 mm) , 1 female (19.5 × 14.8 mm) ( MZUF 2508 View Materials , one chela for genetic analysis as MZUF 4866 View Materials ), Mozambique: Inhaca I., Saco da Inhaca , colls. S. Cannicci & F. Porri, July 1997 . Seychelles: 1 male (16.9 × 13.3 mm) ( MZUF 2980 View Materials ), Seychelles : Mahé, coll. S. Fratini, December 2005 ; 2 males (25.6 × 19.5 mm, 20.0 × 15.9 mm) ( MZUF 2934 View Materials ), Mahé, Port Launay , coll. S. Fratini, December 2005 ; 1 female (17.0 × 13.1 mm) ( MZUF 2937 View Materials ), Mahé, Port Launay , coll. S. Fratini, December 2005 . Rodrigues I.: 1 male (22.0 × 17.3 mm) , 1 female (19.0 × 14.4 mm) ( MZUF 4934 View Materials ), Baie Topaze , coll. M. Vannini, 9 July 1989 ; 1 male (19.1 × 15.2 mm), 2 females (28.0 × 21.5 mm, 23.6 × 18.0 mm) ( MZUF 5004 View Materials ), Baie au Huitres , coll. M. Vannini, 8 July 1989 . Mauritius I.: 2 females (24.5 × 18.8 mm, 18.7 × 13.8 mm) ( MZUF 5005 View Materials ), Melville, coll. M. Vannini, 1 July 1989 . Aldabra : 2 males (23.3 × 18.9 mm, 25.6 × 19.8 mm), 1 female (24.2 × 17.8 mm) ( MZUF 5006 View Materials ), lagoon behind the headquarter, coll. M. Vannini, March 1979 . Iran: 1 male (30.8 × 25.5 mm) , 1 female (30.2 × 24.4 mm) ( MZUF 5014 View Materials ), Bandar Abbas , behind the airport, coll. A. Shahdadi, 10 April 2013 .
Comparative material examined. Metopograpsus thukuhar ( Owen, 1839) : Neotype (here designated), male (32.5 × 27.1 mm) ( ZRC 2020.0219 View Materials ; Figs. 1B View Fig , 2B View Fig , 3B View Fig , 4B View Fig , 5B View Fig ), Hawaii: Oahu, Kewalo, on sea wall, surf zone, Ala Moana area ,
Wakiki, coll. P.K.L. Ng & S.H. Tan, 22 January 2000; 3 males (21.3 × 17.0 mm, 20.5 × 16.4 mm, 17.5 × 13.7 mm), 1 female (28.6 × 23.6 mm) ( ZRC 2000.0405 View Materials ), Hawaii : Oahu , Kewalo , on sea wall, surf zone, Ala Moana area, Waikiki, coll. P.K.L. Ng & S.H. Tan , 22 January 2000; 1 male (20.3 × 16.4 mm) ( NHM 1948.5.3.1), Sandwich Islands (= Hawaii): further collection data unknown ; 1 male (22.1 × 18.5 mm), 2 females (21.8 × 17.6 mm; 20.5 × 16.9 mm) ( NHM 1883.22 ), presented by the Hawaiian Government from the International Fisheries Exposition, 1883 ; 2 males (27.6 × 22.5 mm; 18.0 × 14.2 mm), 3 females (ovigerous: 22.6 × 17.9 mm; 17.2 × 13.1 mm; 13.4 × 10.8 mm) ( NHM 1884.31 ), Hilo, coll. HMS Challenger ; 1 female (ovigerous: 22.6 × 16.7 mm) (NHM not registered), Samoa: Upolu I., coll. Buxton & Hopkins, further collection data unknown ; 1 male (22.0 × 17.9 mm) (NHM not registered), collection data unknown; 2 males (17.6 × 13.1 mm; 13.8 × 10.2 mm), 2 females (ovigerous: 19.4 × 15.00 mm; 15.2 × 11.9 mm) ( NHM 1884.31 ), French Polynesia: Tahiti , near the reefs, coll. HMS Challenger, collection data unknown ; 2 males (22.2 × 17.5 mm; 21.6 × 17.8 mm), 2 females (ovigerous: 23.0 × 18.2 mm; ovigerous: 15.1 × 10.9 mm; 12.5 × 9.5 mm) ( NHM 1884.31 ), Tahiti , Papeete, coll. HMS Challenger, further collection data unknown ; 1 male (13.5 × 10.0 mm) ( NHM 1884.31 ), Fiji: Kandavu, coll. HMS Challenger ; 1 male (18.5 × 15.2 mm) (MNHN-IU-2011-9039), Marquesas Islands :
Nuku Hiva, Hakatea Cove, 8°56′6″S, 140°10′6.006″E, coll. Agence des Aires Marines Protégées, 28 January 2012. Metopograpsus eydouxi H. Milne Edwards, 1853: 1 male (20.2 × 15.2 mm) (MNHN-IU-2000-3547; Fig. 6 View Fig ), Sandwich Islands (= Hawaii), designated as lectotype by Ng et al. (2019: 114); 1 male (25.1 × 19.6 mm), 1 female (21.0 × 16.2 mm) (MNHN-IU-2000-3551, not types ( Ng et al., 2019: 114)), Sandwich Islands (= Hawaii). Metopograpsus intermedius H. Milne Edwards, 1853: 1 male (23.1 × 16.8 mm) (MNHN- IU-2014-11210), unknown locality, designated as lectotype by Tweedie (1954: 128). Pachygrapsus parallelus Randall, 1840 : Syntypes: 2 males (32.7 × 28.0 mm, 11.1 × 8.0 mm), 3 females (26.2 × 22.0 mm, 23.1 × 18.1 mm, 22.0 × 15.0 mm) ( ANSP CA 3545), Sandwich Islands (= Hawaii), colls. T. Nuttall & J.K. Townsend, before 1839.
Diagnosis. Carapace flat, quadrangular, slightly broader than long; branchial groove distinct, branchial region with distinct oblique ridges, urogastric grooves distinct, cardiac and intestinal regions smooth, without any ridge or tubercle. Lateral margins of the carapace entire. Front broad, more than half of greatest width of carapace, deflexed with rugose surface and crenulated free margin, 4 depressed post-frontal lobes along line of frontal deflexion. Suborbital tooth triangular, keeled from tip to base. Exposed surface of base of antenna densely pubescent. Chelipeds subequal, without obvious handedness, fingers stout with spooned tip. Ambulatory legs compressed and with broad merus, dorsal and ventral ridges of 3 distal joints bearing bristles, dactylus with spines. Third and fourth pereiopods with pubescence on lower border of base and ischium. Male pleon with 6 distinct segments, basal one as broad as thoracic sternum, somite 6 rectangular, telson triangular. G1 slender, straight, with twisted ridge along longitudinal axis, apical corneous process elongate.
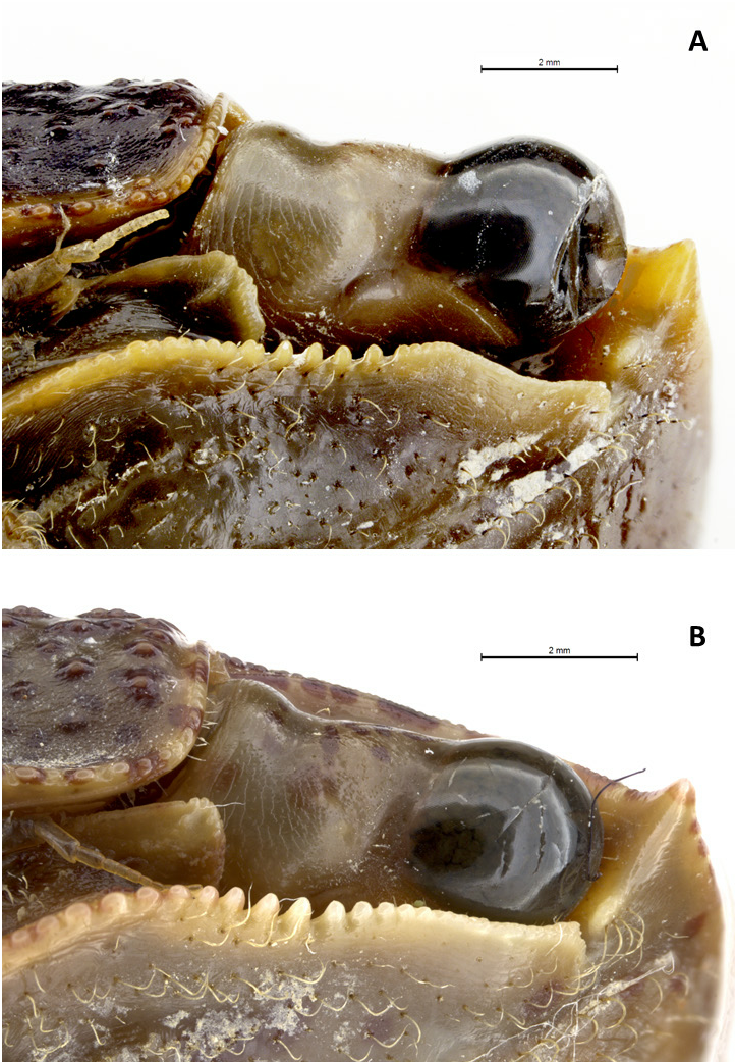
Description. Small- to medium-sized crab (maximum observed CW 34.5 mm). Carapace almost quadrangular, slightly broader than long, greatest width just behind exorbital angles; surface smooth, with transverse ridges lined in rows on branchial region, urogastric grooves evident ( Fig. 1A View Fig ). Front 0.6 (± 0.02) times carapace width (N=6), quite deflexed, slightly concave medially. Post-frontal lobes evident, median ones slightly smaller than lateral ones in width and separated by furrow. Dorsal carapace regions moderately indicated, hepatic region demarcated, cardiac region separated from urogastric region, lateral branchial ridges evident, mid upper orbital border eyelet-shaped, lower orbital border granulate, anterolateral margin with sharp exorbital tooth, lateral margin weakly concave. Suborbital tooth triangular, distally crenulated, with central keel ( Fig. 2A View Fig ).
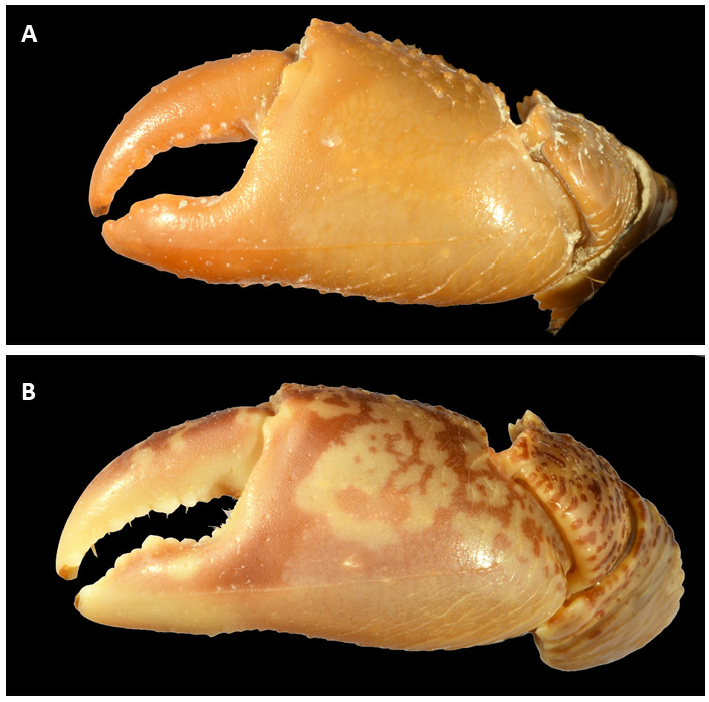
Male chelipeds subequal (51% with right enlarged; 49% with left enlarged, N=10) ( Fig. 3A View Fig ). Chela large (palm length 0.68 ± 0.05 times CW, N= 3 males), robust (length/width ratio = 0.53 ± 0.02, N= 3 males). Merus with grooves on dorsal border and with spines on inner (3 or 4) and upper (4 or 5) borders; ventral border crenulate; inner upper border of carpus with 3 small spines. Upper surface of palm in males with 3 or 4 transverse granulated lines; outer surface of palm smooth, along ventral border a hardly evident ridge crossing several (6 or 7) transverse ridges, ventral border coarsely granular until outer surface of fixed finger, centrally gently hollowed; inner surface of palm coarsely smooth except area facing carpus; length of cutting edge of fixed finger 0.39 ± 0.04 (N= 3 males) times maximum propodus length. Dactylus relatively stout, slightly curving in dorsal view, length 0.57 ± 0.05 (N=3) times maximum propodus length. Fingers with chitinous spoon-shaped tips, with gape when closed. Cutting edge of both fingers with series of inconspicuous teeth. Female chelipeds smaller than in males, subequal as in males, ratio of palm length to carapace width ca. 0.60 in female paratype ( MZUF 5008 View Materials ). Merus and carpus as in males, upper surface of palm with 3 or 4 transverse and sparse granules, inferior border of palm with line along its length and 4 lines of not relevant transverse ridges, both fingers with chitinous spoon-shaped tips .
Ambulatory legs relatively long, with broad merus (length/ width ratio = 0.46 ± 0.01, N= 11 males), flattened; fourth and fifth pereiopods longest ( Fig. 1A View Fig ). Edges of last three joints bearing bristles, dactylus with spines. Lower border of third and fourth base and ischium pubescent.
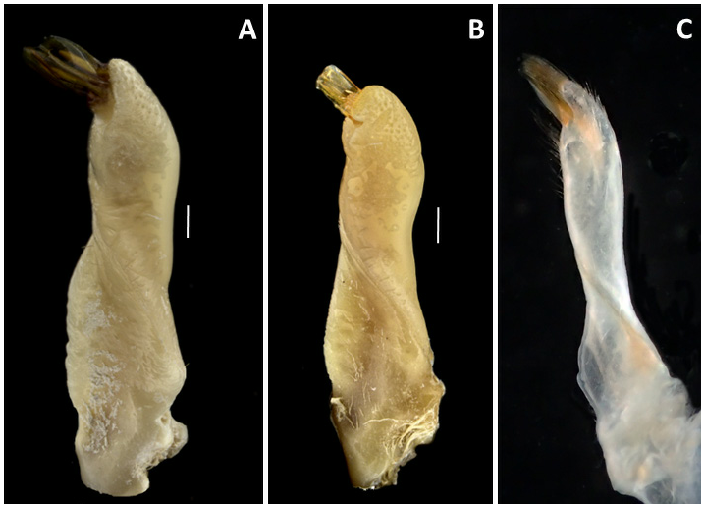
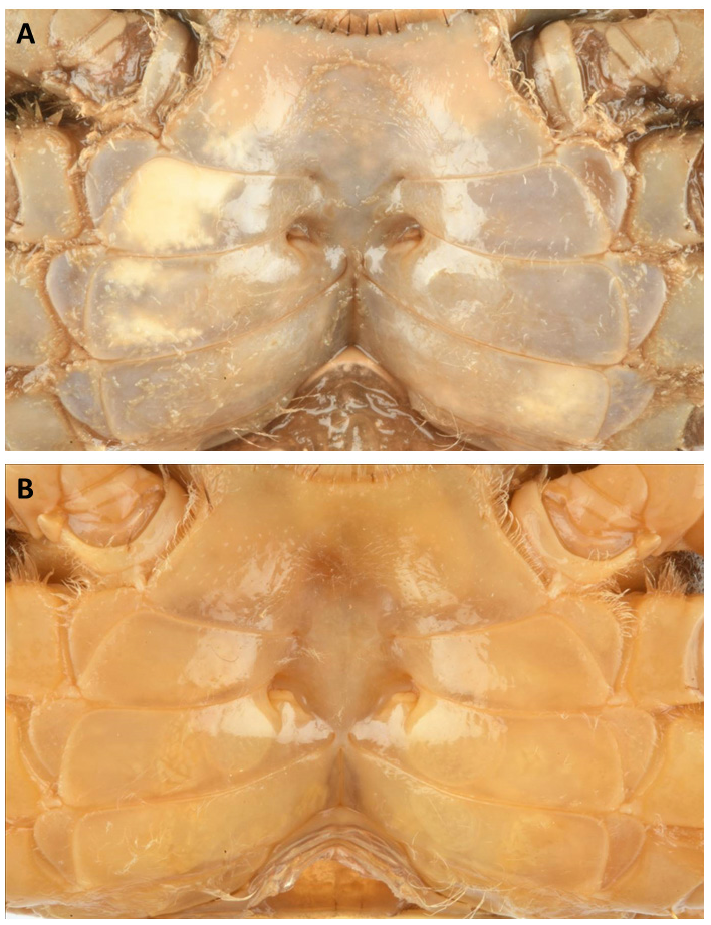
Male pleon triangular; telson triangular and slightly longer than somite 6. Somite 6, 1.60 ± 0.04 (N=3) times wider than the general shape of the G 1 in M. thukuhar is distally more swollen, with a rounded hump and a shorter corneous tip ( Fig. 4B View Fig ). However, only fully adult specimens can be distinguished by their G1, as in males with CW <23 mm such differences are not always marked (see Fig. 4C View Fig ). Adult females of M. cannicci , new species, have a trapezoidal sternal vulvar cover ( Fig. 5A View Fig ), while in female M. thukuhar the cover is wider and higher ( Fig. 5B View Fig ). The live colouration is also very variable between and within both species, thus not helping in possible identification. It is hypothesised that it probably depends upon the background on which the crabs thrive.
long. Somites 4 and 5 trapezoidal. Somite 3 widest, laterally convex. Somite 2 following lateral edges. G1 slender, straight, with twisted ridge along its axis, distal part slender, apical corneous process elongate ( Fig. 4A View Fig ). Female pleon fringed with long setae, broad, evenly rounded, in adults laterally touching coxae of walking legs; telson half-moon shaped, ca. 0.4 times wider than long. Vulva in depression at the border of sternite 6 with sternite 5, operculum on inner part with an evident trapezoidal sternal vulvar cover in adult females ( Fig. 5A View Fig ).
Colour. In life, crabs are mottled brown-green and cream, and distinctly glossy ( Fig. 7 View Fig ). Crabs with CW> 24 mm are purple on the outer face of the chelar palm. Preserved specimens have brown spots, still visible on a creamy patterned carapace, cheliped cream-coloured with brownish-orange tips.
Etymology. The new species is dedicated to our colleague and friend, Dr Stefano Cannicci, who devoted his research career to study mangrove fauna, especially of East Africa; the name here is used as a noun in apposition.
Remarks. The description of M. cannicci , new species, is quite challenging as, at first sight, all the examined specimens from the Red Sea to the central Pacific Ocean are morphologically very similar. However, the G1 of M. cannicci , new species, is slightly different in shape from that of M. thukuhar , being more slender and less inflated at its distal end, with a more acute hump ( Fig. 4A View Fig ). Moreover, Owen’s (1839) original description of Grapsus thukuhar , based on a single specimen (without sex specification) collected from Oahu (Hawaii), provided diagnostic characters that are very common for all species of Metopograpsus : “ Grapsus clypei lateribus striatis; humeris, ulnis, carpisque interne spinosis; femoribus supra et subtus spinosis […] Color, fulvus brunneo-punctatissimus. Carapace quadrilateral, broadest in front; the sides slightly converging to the posterior angles, which are truncated. Rostrum very broad, inclined, supporting four prominences, of which the laterals are the largest. No teeth at the sides of the shell, but the anterior angles produced and acute; oblique lines over the branchial regions. Chelae equal, short, obtuse; humeri with two spines; carpi with one or two spines internally; manus slightly tuberculated at the upper part, the remainder smooth and mottled with purple. Claws compressed, the femora with two or three spines at their apices, the other joints hairy, the terminal ones armed with short brown spines, sprinkled over with minute brown spots, like the skin of Sepia officinalis ” ( Owen, 1839: 80) . Unfortunately, the holotype was destroyed during the 1941 bombing of the Royal College of Surgeons (London, UK), where it was deposited (Sarah Pearson, personal communication).
Banerjee (1960: 174) reported the following diagnostic characters for M. thukuhar : “lateral margins of the carapace scarcely convergent backwards, upper surface of the base of the antenna densely pubescent, 6th segment of the male abdomen equal to or slightly longer than fifth, compressed finger-shaped chitinous projection present at the top of the male pleopod”. He also mentioned a blunt sub-orbital tooth, not keeled from tip to base, but this character was found to be extremely variable in the examined specimens, finding in many of them a central keel, as already pointed out by Vannini & Valmori (1981) in their remarks on specimens of M. thukuhar from Somalia. The specimens from Hawaii have an evident keel in the suborbital tooth too ( Fig. 2B View Fig ).
The male specimen from ZRC ( Figs. 1B View Fig , 2B View Fig , 3B View Fig , 4B View Fig , 5B View Fig ) is very similar to the specimen figured by Owen (1839: pl. 24, figs. 3, 3a), and it was collected from the same locality. Therefore, we decided to designate it as the neotype of M. thukuhar to stabilise its taxonomy, and to serve as the primary point for comparison with M. cannicci , new species.
Three other names have been treated as synonyms of Metopograpsus thukuhar (Owen) by previous workers (e.g., Tweedie, 1949, 1954; Banerjee, 1960; Davie, 2002; Ng et al., 2008), viz. Metopograpsus eydouxi H. Milne Edwards, 1853 (type locality: Hawaii), Metopograpsus intermedius H. Milne Edwards, 1853 (type locality: unknown), and Pachygrapsus parallelus Randall, 1840 (type locality: Hawaii), and their type material have also been examined in this work. The description of Pachygrapsus parallelus by Randall (1840) is based on the external shape of the carapace, claws and legs with no mention of gonopods. The lectotype of M. eydouxi was selected and figured recently by Ng et al. (2019: 114, fig. 6F). The G1s of M. eydouxi and P. parallelus could not be observed, however, as the specimens have been dried and are very fragile. As for Metopograpsus intermedius H. Milne Edwards, 1853 , Ng et al. (2019: 115, fig. 7B) reconstructed the intricate history of the type specimen as follows: Tweedie (1949) indicated that Metopograpsus intermedius H. Milne Edwards, 1853 , was represented in the collection of the MNHN by two syntypes belonging to two distinct species; the smaller of them being a M. thukuhar , whereas the larger agreed with the description of M. gracilipes De Man, 1891 , according to the examination of the G1 sketches by Isabella Gordon. Tweedie (1954) designated the smaller syntype as the lectotype of M. intermedius , in order to define M. intermedius as a junior synonym of M. thukuhar and to establish M. gracilipes (presently a synonym of M. frontalis Miers, 1880 ) as the valid name for the species represented by the larger co-type of M. intermedius . Figure 6 View Fig shows the neotype of M. thukuhar and compares it with Owen’s figure of M. thukuhar , and photographs of the types of M. intermedius , M. eydouxi , and P. parallelus .
Ecology. Within its geographical range, M. cannicci , new species, is often associated with mangrove forests. Among the specimens analysed in this study, the Red Sea crabs were collected among the pneumatophores of Avicennia marina shrubs, while all the other specimens were collected in marine creeks and fringing mangroves ( Vannini & Valmori, 1981; Fratini et al., 2000). However, populations of M. cannicci are also known to colonise man-made structures in sheltered areas, such as harbours and vertical jetties, where they can be abundant on wooden and concrete vertical surfaces. In East Africa, M. cannicci colonises both the seaward fringe, where it can be found in sympatry with M. messor and M. (2018), now providing morphological and nuclear DNA evidences that the former species Metopograpsus thukuhar consists of a complex of two pseudocryptic species, one of which is here described as Metopograpsus cannicci , new species.
The term pseudocryptic is hereby used in the sense of Ragionieri et al. (2012) and Fratini et al. (2019), i.e., minor morphological differences discovered, after recognising the genetic distinctness. With M. cannicci , new species, the number of species belonging to Metopograpsus increases to seven. Since the type specimen used by Owen (1839) for the description of M. thukuhar was from Hawaii, the original name remains associated to populations occurring in the eastern Indian and Pacific oceans, while M. cannicci , new species, refers to populations from the Red Sea, East Africa, western Indian Ocean islands, and the Persian Gulf. oceanicus , and the denser part of the Rhizophora mucronata forests, where it is usually the only species of Metopograpsus present (see Cannicci et al., 1996). This species is known to be a generalist feeder and a very fast and agile predator of fiddler and sand bubbler crabs ( Fratini et al., 2000).
Distribution. Red Sea, East African coast (from Somalia ( Vannini & Valmori, 1981, quoted as M. thukuhar ) to Mozambique), Seychelles; Dar es Salaam ( Hartnoll, 1975, quoted as M. thukuhar ); Madagascar, Toliara and Toamasina ( Crosnier, 1965, quoted as M. thukuhar ); Mauritius ( Michel, 1964, quoted as M. thukuhar ; present paper); Iran ( Naderloo & Türkay, 2012; Naderloo, 2017, in both papers quoted as M. thukuhar ; present paper).
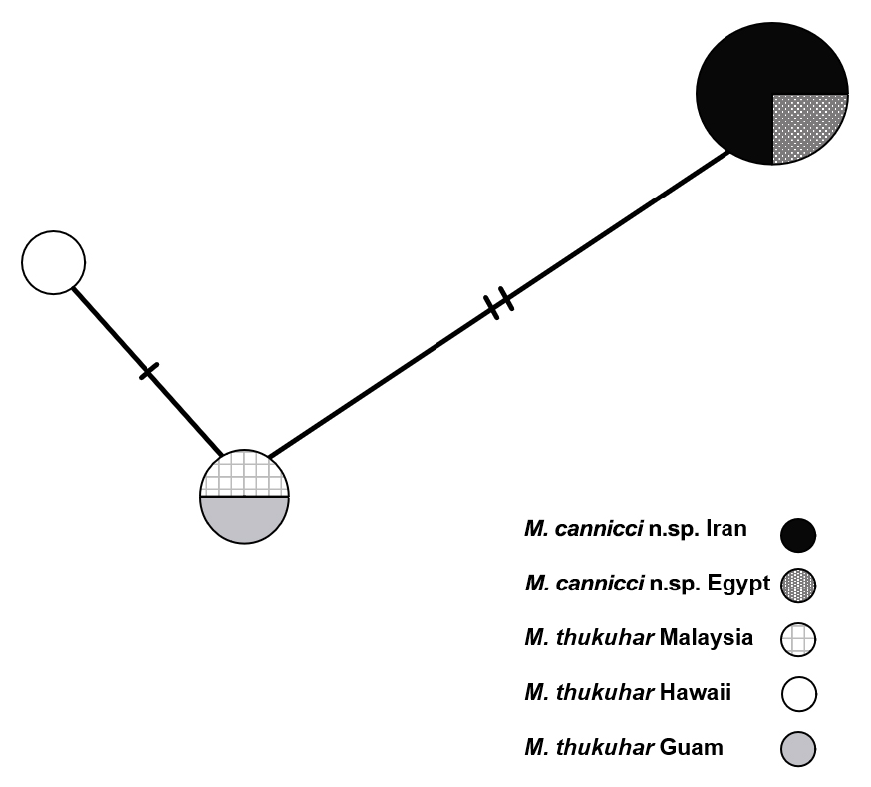
Nuclear genetic evidence. The TCS genotype network ( Fig. 8 View Fig ) shows the clustering of seven individuals of Metopograpsus , formerly treated as M. thukuhar , across three individual 28S genotypes. A genotype is here defined as a specific sequence of nucleotides within the 28S nuclear gene region and is visualised as a circle in Figure 8 View Fig . Short transverse lines indicate the number of mutations separating the different genotypes. The most common genotype is shared by the four individuals from Iran and Egypt and is separated by two mutations from a second genotype comprising specimens from Guam and Malaysia. A third genotype is separated by one additional mutation from the latter ones and was found in a sample from Hawaii.



| NHM |
University of Nottingham |
| ANSP |
Academy of Natural Sciences of Philadelphia |
| CA |
Chicago Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.