
Spinoncaea, 2003
|
publication ID |
https://doi.org/10.1046/j.1096-3642.2003.00056.x |
|
DOI |
https://doi.org/10.5281/zenodo.5110304 |
|
persistent identifier |
https://treatment.plazi.org/id/77650E13-FFF3-3604-FECA-FC224872755B |
|
treatment provided by |
Carolina |
|
scientific name |
Spinoncaea |
| status |
sp. nov. |
SPINONCAEA TENUIS SP. NOV.
Synonymy
None.
Type locality
Northernmost Red Sea, 27∞25.00¢N, 34∞04.98¢E: Stn. 156; R/V Meteor leg 44/2: collected 4 March 1999 with MSN 0.055 mm net (Haul 9/3); depth 300–350 m; total water depth 798 m
.
Material examined
See Table 3 View Table 3 (c).
Preliminary note
The typical form of S. tenuis occurring in the Red Sea proper is described in detail in the following. Morphological differences of a more elongate female form that was found in the Strait of Bab al Mandab (southern entrance of the Red Sea) and in various other geographical localities will be given below under ‘ Variability ’.
Description of female
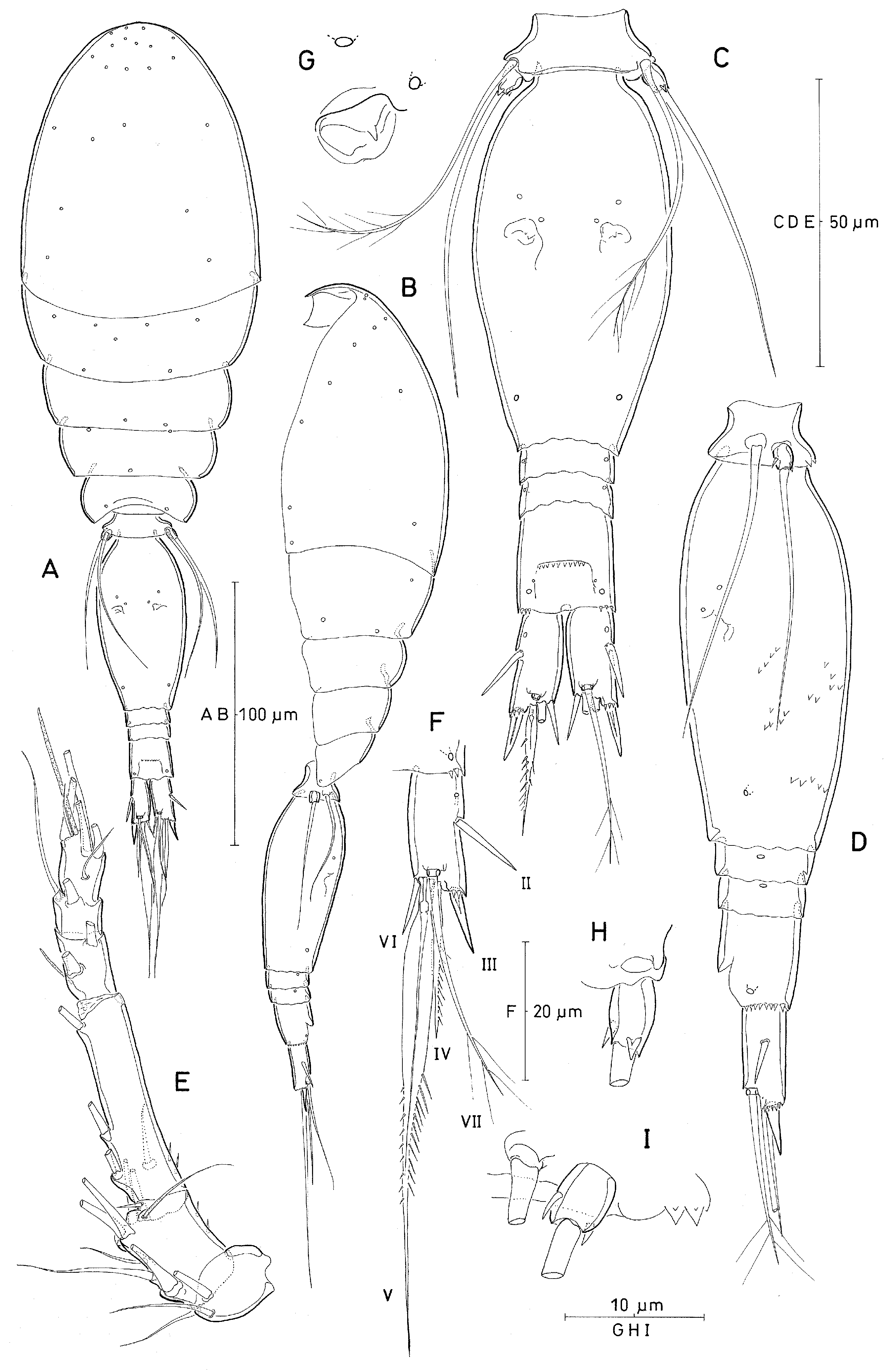
Body length (measured in lateral aspect; from anterior margin of rostral area to posterior margin of caudal rami, calculated as sum of individual somites): 370 Mm [traditional method: 290 Mm (range: 280–300 Mm), based on 2 specimens].
Exoskeleton moderately well chitinized. Prosome 2.1 times length of urosome, excluding caudal rami, 1.8 times urosome length including caudal rami. P2-bearing somite without dorso-posterior projection in lateral aspect ( Fig. 12B View Figure 12 ). Integumental pores on prosome as indicated in Fig. 12 View Figure 12 (A,B). Pleural areas of P4-bearing somite with rounded posterolateral corners.
Proportional lengths (%) of urosomites are 8.2: 62.7: 5.9: 5.9: 17.3. Proportional lengths (%) of urosomites and caudal rami are 7.2: 54.6: 5.1: 5.1: 15.0: 13.0.
P5-bearing somite with 2 paired midventral spinous processes ( Fig. 12I View Figure 12 ).
Genital double-somite oval-elongate, 2.0 times as long as maximum width (measured in dorsal aspect) and 2.1 times as long as postgenital somites combined; lateral margins rounded, largest width measured at about halfway the distance between anterior and posterior margin, posterior part tapering. Pore pattern on dorsal surface as in Fig. 12 View Figure 12 (C); ventral surface with several rows of minute spinules as indicated in Fig. 12 View Figure 12 (D). Paired genital apertures located dorsally at about half distance from anterior margin of genital double-somite; armature difficult to discern, possibly represented by 1 diminutive spinule ( Fig. 12G View Figure 12 ). Hyaline frill on posterior margin of genital double-somite and postgenital somites weakly undulate.
Anal somite about as wide as long; slightly shorter than CR (measured along outer margin) or slightly longer than CR (measured along inner margin) ( Fig. 12F View Figure 12 ), variation in length to width ratio not examined. Dorsal surface with two pairs of secretory pores near posterior margin, paired dorsal sensillae anterior to anal opening absent or not discernible. Anterior margin of anal opening (vestigial anal opening) with minute spinules. Posterior margin of somite finely serrate ventrally and laterally ( Fig. 12C,D View Figure 12 ).
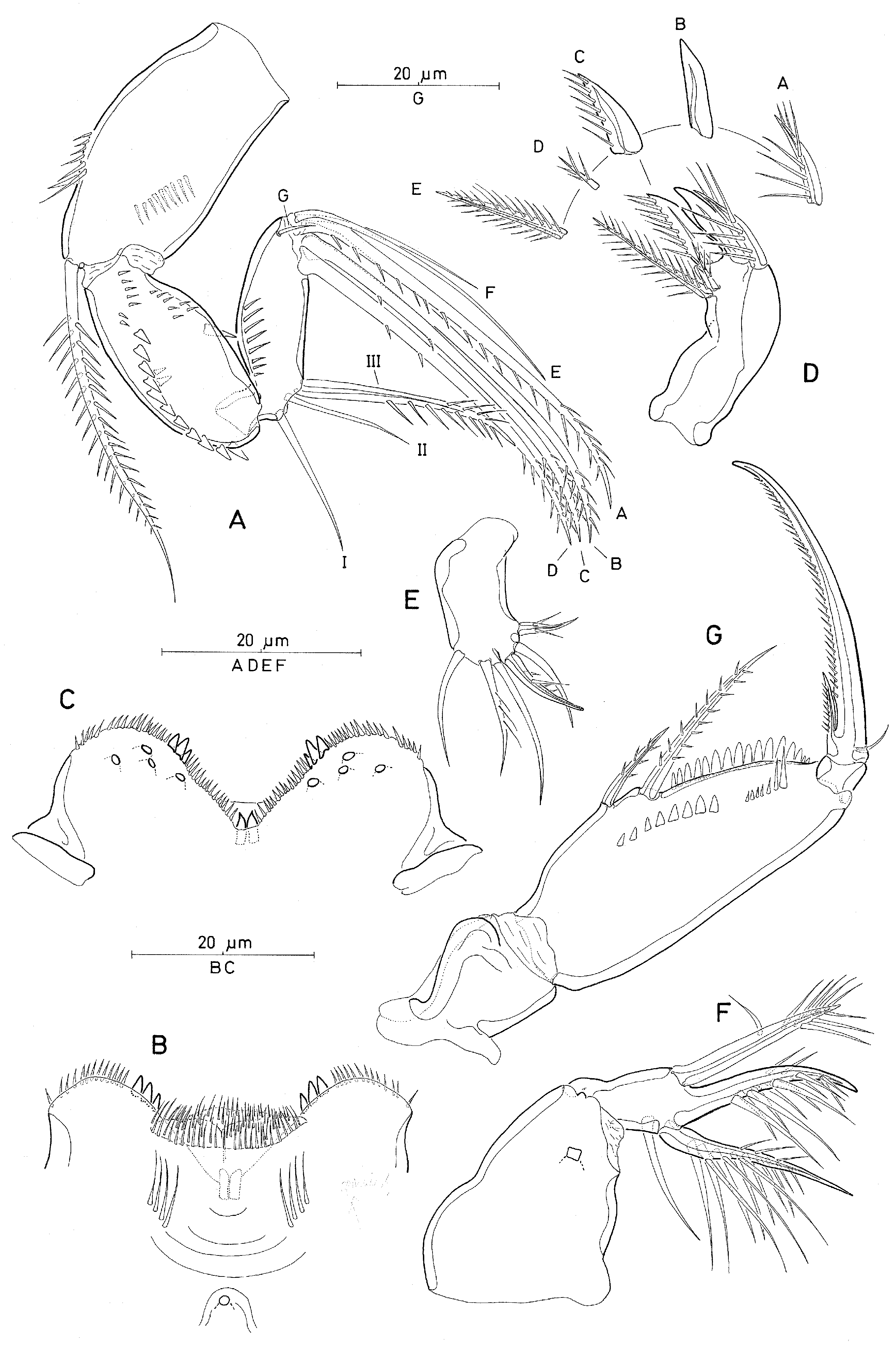
Caudal ramus ( Fig. 12F View Figure 12 ) 1.9 times longer than wide measured along inner margin and 2.3 times longer than wide measured along outer margin; variation in length to width ratio not examined. Armature consisting of 6 elements, numbered by Roman numerals in Fig. 12 View Figure 12 (F): seta II small, spiniform, and unornamented; seta III strong, spiniform and unornamented, base of seta concealed by serrate margin of CR ( Fig. 12D,F View Figure 12 ); seta IV about twice as long as seta III, moderately lanceolate and bipinnate ( Fig. 12C View Figure 12 ); seta V about 3 times longer than seta IV, lanceolate, naked at anterior half, posterior half with short spinules along inner margin and longer pinnules along outer margin [this seta easily gets lost during handling]; seta VI shorter than seta III, unornamented; seta VII long, about half length of seta V, bipinnate at distal part and bi-articulate at base. Inner margin of CR unornamented. Dorsal anterior surface ( Fig. 12F View Figure 12 ) with secretory pore near insertion of seta II.
Antennule 6-segmented ( Fig. 12E View Figure 12 ), relative lengths (%) of segments measured along posterior nonsetiferous margin 8.0: 17.8: 41.7: 14.7: 7.4: 10.4. Armature formula: 1-[3], 2-[8], 3-[5], 4-[2+ae], 5-[2 (ae not discernible)], 6-[5+(1+ae)]. Aesthetasc on segment 4 very slender, no aesthetasc discernible on segment 5 [but see male, Fig. 15 View Figure 15 (G)]; apical aesthetasc well developed and fused basally to adjacent seta. Segment 2 and proximal part of segment 3 ornamented with few, short pinnules along inner, nonsetiferous margin.
Antenna 3-segmented, distinctly reflexed ( Fig. 13A View Figure 13 ). Coxobasis with row of spinules along outer and inner margins and with pattern of denticles on outer (exopodal) margin; with long seta at inner distal corner, ornamented unilaterally with long spinules, subdistal one very long. Endopod segments about equal in length; proximal endopod segment elongate-oval, expanded outer margin bearing short spinular row; posterior surface with row of short, strong denticles along inner margin. Distal endopod segment about 4 times longer than wide, with narrow cylindrical base articulating with the proximal endopod segment; posterior surface with row of short spinules along outer margin; lateral armature consisting of 2 bare setae, with seta II shorter than seta I, and 1 long spiniform seta (III), ornamented with strong spinules bilaterally at distal half and unilaterally at proximal part, seta IV absent; distal armature consisting of 4 long spiniform setae (A–D), ornamented with spinules bilaterally at distal part and unilaterally along entire (A) or half (C,D) length, 3 naked setae (E–G) of varying length, posterior seta (G) reduced to small spinule.
Labrum ( Fig. 13B,C View Figure 13 ) distinctly bilobed. Distal (ventral) margin of each lobe with 3 marginal teeth medially, differing slightly in size, long row of very small spinules at outer ventral margin and row of small spinules or denticles along inner margin. Median concavity covered anteriorly by overlapping rows of fine spinules. Anterior surface ( Fig. 13B View Figure 13 ) with 2 paired rows of long setules; median swelling weakly developed, with large secretory pore proximally. Posterior wall of medial concavity with two chitinized spinous teeth, flanked by row of minute denticles or spinules ( Fig. 13C View Figure 13 ). Posterior face with 3 paired secretory pores located distally on each lobe.
Mandible ( Fig. 13D View Figure 13 ) gnathobase with 4 elements: 2 setae and 2 blades, for indication of elements see Fig. 3 View Figure 3 (D). Ventral element (A) as long as ventral blade (B), with long, fine setules along dorsal side; ventral blade strong and spiniform, unornamented; dorsal blade (C) strong and broad, spinulose along entire dorsal margin; dorsal element D absent, element E setiform and bipinnate.
Maxillule ( Fig. 13E View Figure 13 ) indistinctly bilobed, surface ornamentation not discernible. Inner lobe (praecoxal arthrite) with 3 elements: outermost element spiniform, swollen at base, ornamented with 2 rows of spinules, tip possibly with tubular extension; middle element setiform and unipinnate; innermost element smallest, located along concave inner margin close to other elements, ornamented with few spinules. Outer lobe with 3 elements [innermost element absent]; outermost element curved and bare, similar in length to the following; element next innermost with short spinules unilaterally; innermost seta longest and unipinnate.
Maxilla ( Fig. 13F View Figure 13 ) 2-segmented, allobasis nearly as long as syncoxa. Syncoxa unarmed, surface ornamented with 1 large secretory pore. Allobasis produced distally into slightly curved claw bearing 2 rows of strong spinules along medial margin; spinules of inner row shorter, proximal group of 2 spinules on outer row longer, small gap between proximal and distal group; outer margin with strong seta extending to tip of allobasal claw, ornamented with long spinules bilaterally at distal part and 1 long spinule subdistally; inner margin with slender naked seta and strong basally swollen spine with double row of long spinules along the medial margin and few spinules along outer margin ( Fig. 13F View Figure 13 ).
Maxilliped ( Fig. 13G View Figure 13 ) 4-segmented, comprised of syncoxa, basis and 2-segmented endopod. Syncoxa unarmed, surface ornamentation not discerned. Basis elongate, palmar margin with 2 spiniform elements, proximal element bare, about half the length of distal one, which is bipinnate; fringe of short spatulated spinules between proximal seta and articulation with endopod; anterior surface with row of broad spatulated pinnules and short spinules of varying length along palmar margin as illustrated in Fig. 13 View Figure 13 (G). Proximal endopod segment unarmed. Distal endopod segment drawn out into long curved claw, with pinnules along entire concave margin; accessory armature consisting of minute, naked seta on outer proximal margin and unipectinate spine fused basally to inner proximal corner of claw.
Swimming legs 1–4 biramous ( Fig. 14A–D View Figure 14 ), with 3- segmented rami. Spine and setae formula as for S. ivlevi . Intercoxal sclerite of P1 ornamented with paired row of long, fine setules ( Fig. 14A View Figure 14 ). Coxae and bases with sparse surface ornamentation as figured. Bases with naked outer seta, arising from posterior surface, longest in P4; inner portion of basis slightly produced adaxially into rounded (P1) or pointed (P2– P4) process, bearing short spinule(s) along inner margin in P1 ( Fig. 14A View Figure 14 ), pointed process reduced in P4. Inner basal seta on P1 spiniform and naked. Respective legs without distinct length differences between exopod and endopod. Bases of spines on exopodal and endopodal segments anteriorly surrounded by small spinules, which are hardly discernible. Surface ornamentation of all segments sparse.
Exopods. Outer margin of exopod segments with well developed serrated hyaline lamella; inner margin of proximal exopod segments with long setules. Secretory pore present on posterior surface of distal segments in P2–P4, also present on anterior surface of P2. Hyaline lamellae on outer spines well developed; outer and terminal spines of P1 with subapical tubular extension, except for spine on exp-2 and proximalmost spine on exp-3. Spine on middle segment and proximal spine on distal segment of P2–P3 reduced in length. Terminal spine slightly longer than (P1) or equal in length to (P2–P4) distal exopod segment.
Endopods. Outer margin of endopod segments with fringe of long setules, except for outer margin of P4 enp-1 naked and enp-2 ornamented with 1 or 2 spinule(s), variable within and between specimens examined [additional spinules possibly indicated by insertion on margin (?), stippled in Fig. 14 View Figure 14 (D)]. Inner seta of proximal endopod segment short, slightly swollen and ornamented with spinules bilaterally (P1-P3) or spiniform with strong spinules bilaterally (P4). Inner setae of P4 enp-2 and -3 reduced in length, in particular those on enp-2, with proximal seta reaching little further than insertion of distal seta and distal seta not reaching half length of seta on distal segment. Distal endopod segments with large secretory pore on anterior face in P1. Inner margin of P1 enp-2 ornamented with 2–3 long spinules. Distal margin of P1 enp-1-and -2 ornamented with row of short denticles or spinules on anterior face; outer margin of P1 enp-3 terminating in long process obscuring insertion of distalmost inner seta. Distal margin of P2–P3 not produced into conical process but apical pore of reduced process present, located laterally between subdistal and distal spine ( Fig. 14B,C View Figure 14 ). Outer subdistal spine on P3 equal in length to outer distal spine, reaching as far as insertion of this spine. Inner setae of distal endopod segments in P2–P4 with spinule comb along proximal inner margin; this comb not obvious in P1; also present on distal inner seta of middle endopod segment in P3–P4.
P5 ( Fig. 12H,I View Figure 12 ) comprised of very long, sparsely plumose seta arising from lateral surface of somite, and small free segment representing exopod. Exopod 1.5 times longer than wide, bearing single long, bare seta; posterior margin ornamented with small spinule dorsally ( Fig. 12I View Figure 12 ) and small spinous process ventrolaterally ( Fig. 12H View Figure 12 ).
P6 ( Fig. 12G View Figure 12 ) represented by operculum closing off each genital aperture; possibly armed with a short spinule as indicated, variable in conspicuousness [not seen in specimens from Adriatic Sea].
Egg-sacs not observed.
Description of male
Body length: 356 Mm [traditional method: 294 Mm, range: 280–300 Mm, based on 10 specimens]. Pore pattern on prosome not fully discerned ( Fig. 15A View Figure 15 ). Sexual dimorphism in antennule, maxilliped, P6, and in genital segmentation, slight modification in length of seta on P5.
Proportional lengths (%) of urosomites (excluding caudal rami) 8.5: 65.5: 3.1: 3.6: 3.6: 15.7; proportional lengths (%) of urosomites (caudal rami included) 7.4: 57.1: 2.7: 3.1: 3.1: 13.7: 12.9. Posterior margin of leg 5-bearing somite with paired row of 3 midventral spinous processes ( Fig. 15E View Figure 15 ). Genital somite with length to width ratio 1.7: 1.
Caudal rami 1.9 times longer than wide measured along inner margin and 2.3 times longer than wide measured along outer margin, similar to female. Caudal setae with proportional lengths as in female, except for seta III being slightly shorter and seta IV being somewhat longer than in female. Dorsal surface of genital somite covered with paired posterolateral pore ( Fig. 15D View Figure 15 ). Surface of genital flaps ornamented with several rows of strong denticles or spinules; ventral surface of anal segment without ornamentation.
Antennule ( Fig. 15G View Figure 15 ) 4-segmented; distal segment corresponding to fused segments 4–6 of female; relative lengths (%) of segments measured along posterior nonsetiferous margin 9.5: 16.9: 45.9: 27.7. Armature formula: 1-[3], 2-[8], 3-[4], 4-[9+2ae+(1+ae)], aesthetascs very small and slender, apical aesthetasc fused basally to adjacent seta, length not fully discerned (stippled in Fig. 15G View Figure 15 ). Ornamention on inner margin of segments 2 and 3 as in female ( Fig. 15g View Figure 15 ).
Maxilliped ( Fig. 15B,C View Figure 15 ) 3-segmented, comprising syncoxa, basis and 1-segmented endopod. Syncoxa missing in specimen figured. Basis elongate, moderately inflated in proximal half, forming small bulbous swelling; anterior surface with row of short spinules along palmar margin, developed into small distal flap ( Fig. 15B View Figure 15 ); posterior surface with 1–2 rows of short spatulated spinules of graduated length along palmar margin ( Fig. 15B,C View Figure 15 ); with 1 long seta within the longitudinal cleft, corresponding to distal seta in female, ornamented with strong spinules bilaterally; proximal seta absent. Endopod drawn out into long curved claw, concave margin ornamented with pinnules along entire concave margin as in female; accessory armature consisting of short, unipectinate spine basally fused to inner proximal corner of claw; tip of claw without hyaline apex.
P1–P4 with armature and ornamentation as in female, ornamentation of second endopod segment in P4 with several spinules along outer margin (as usually found in oncaeids), not only single spinule as in female.
P5 ( Fig. 15D,F View Figure 15 ) exopod with general shape and armature as in female; exopodal seta and long seta arising from lateral surface of somite shorter than in female.
P6 represented by posterolateral flap closing off genital aperture on either side; covered by pattern of denticles as shown in Fig. 15 View Figure 15 (E); posterolateral corners protruding laterally so that they are well discernible in dorsal aspect ( Fig. 15A,D View Figure 15 ).
Spermatophore oval ( Fig. 15D View Figure 15 ), of variable size according to state of maturity; swelling of spermatophore during development not affecting shape and relative size of genital somite.
Etymology
The specific name Latin tenuis , meaning slender, refers to the smaller seta III on the caudal ramus and to the general habitus of the species.
Remarks
Taxonomy. Spinoncaea tenuis differs from S. ivlevi and S. humesi sp. nov. by the different proportions of the urosomites, with the genital double-somite measuring about 2/3 of the urosome, whereas it is only about half the length of the urosomites in the 2 other species. Further differences separating S. tenuis from its congeners are the length ratio and form of caudal setae, in particular the more slender spinous element III, the greater length and different form of seta IV, being slightly dilated and 2.5 times longer than seta III, whereas it is not dilated and only about 1.5 times longer than seta III in S. ivlevi and S. humesi , and the greater length of seta V, being 3 times longer than seta IV, whereas it is only about 2 times longer than this seta in the two other species. The form and length of seta IV appears to be a useful tool in separating species of Spinoncaea from closely related species, such as O. tregoubovi and O. prendeli , because it was found to be consistent within all species (including their form variants) described during the present study.
From S. humesi the species can be most readily be distinguished by the spine count on the distal exopod segment of P2, showing 3 spines in S. tenuis , but only 2 spines in S. humesi . S. tenuis and S. ivlevi share the same swimming leg armature, but in addition to the characters given above they can be separated by (1) the number of elements on the mandible ( 4 in S. tenuis , 5 in S. ivlevi ), (2) the ornamentation of the coxobasal seta in the antenna (unilaterally, with 1 long subdistal spinule in S. tenuis , bilaterally in S. ivlevi ) and (3) the ornamentation on the anterior surface of the labrum (2 rows of spinules in S. tenuis , one in S. ivlevi ). Another slight difference between the two species is found in the ornamentation of the inner margin of P4 enp-2, showing the typical fringe of several long setules in S. ivlevi , whereas there are 2 or even only 1 long spinule(s) in S. tenuis .
Variability. Female specimens of Spinoncaea tenuis displayed considerable morphological variation both within and between geographical regions. Differences in body morphology observed during the present study included (1) the form of the genital double-somite and (2) the length to width ratio of (a) the genital doublesomite, (b) the anal somite and (c) the caudal rami. Based on these characters, two form variants of S. tenuis were established, a typical form and an elongate form, with the latter generally exhibiting higher ratios ( Table 8). The elongate form of S. tenuis from the Adriatic Sea ( Fig. 16A View Figure 16 ) was examined in more detail and differed from the typical Red Sea form in the following characters: (1) body length: 390 Mm [traditional method: 315 Mm, range: 300–330 Mm, based on 6 specimens from the Adriatic Sea]; (2) proportional lengths (%) of urosomites and caudal rami are 7.1: 52.4: 5.6: 5.6: 14.6: 14.6; (3) genital doublesomite more elongate than in typical form ( Fig. 16B View Figure 16 ), 2.2 times as long as maximum width (measured in dorsal aspect). Genital apertures located somewhat more anterior than in typical form, at 2/5 of distance from anterior margin, armature of genital apertures not clearly discernible; (4) anal somite slightly shorter than CR (measured along outer margin) or about as long as CR (measured along inner margin) ( Fig. 16B View Figure 16 ); (5) caudal ramus ( Fig. 16B View Figure 16 ) longer than in typical form, 2.4 times or 2.6 times longer than wide measured along inner or outer margin, respectively; armature and ornamentation mainly as in typical form, except for seta VII being slightly longer and ornamentation of seta V as figured ( Fig. 16B View Figure 16 ); (6) swimming legs with armature and ornamentation as in typical form, except for basal seta on P4 stronger and much longer, reaching beyond distal exopod spine ( Fig. 16C View Figure 16 ). Reduced inner setae on P4 enp-2 slightly longer than in typical form, with proximal seta almost reaching insertion of distal seta and distal seta reaching half length of seta on distal segment ( Fig. 16C View Figure 16 ). Ornamentation on inner margin of P4 enp-2 similar to typical form, showing only 1 single spinule [insertion of further spinules not present]; (7) P5 with outer basal seta slightly longer than in typical form ( Fig. 16B View Figure 16 ), armature of P6 not discernible, egg-sacs not found. Antennule, antenna and mouthparts similar to typical form.
In the male, morphological differences between specimens of the Red Sea and the Adriatic Sea became less obvious and the assessment of an elongate male morph in the Adriatic Sea appears to be questionable. Adriatic specimens of S. tenuis ( Fig. 16D View Figure 16 ) were similar in size, measuring 340 Mm in body length [traditional method: 296 Mm, range: 290–300 Mm, based on 5 specimens from the Adriatic Sea] and showed similar body proportions and urosome segmentation ( Fig. 16D View Figure 16 ). However, the genital somite of specimens from the Adriatic was slightly longer than in the typical Red Sea form, with a length to width ratio of 1.8: 1, and the caudal rami were longer, being about 3 times longer than wide (2.8 times longer than wide measured along inner margin, 3.0 times longer than wide measured along outer margin, respectively). Proportional lengths of caudal setae were similar to the typical form, except for seta VII, which was slightly longer. Pore patterns on the prosome and the urosome were not fully discerned in males from the Adriatic Sea.
The two forms of S. tenuis differed in geographical distribution ( Table 8): the typical form of S. tenuis occurred in the entire Red Sea and was also present in the northern Arabian Sea. The elongate form occurred in the Mediterranean Sea and in the NW Pacific (Kuroshio Extension). Specimens from the NE Pacific (Monterey) were tentatively assigned to the elongate form of S. tenuis on the basis of caudal rami characters, but they showed intermediate values between the two forms in the other characters examined ( Table 8). The observed morphological differences between the two female forms of S. tenuis – although being geographically separated – were not regarded as sufficient to warrant recognition of the two forms as separate species, and the Red Sea specimens of S. tenuis are regarded as conspecific with those from the Mediterranean. Specimens from the North Pacific are regarded as conspecific as well, but need to be examined in more detail, as their identification in the present study was based on a limited number of characters only. Based on the present preliminary data, a wide geographical distribution of the elongate form(s) of S. tenuis is indicated, whereas the typical form appears to be restricted to the Arabian and Red Sea.
Two additional morphs of the tenuis - type were found outside the Red Sea main basin during the present study (cf. Table 3 View Table 3 ): In the Adriatic Sea, a single female specimen was separated (= Spinoncaea sp. A), which differed in the form of the genital doublesomite, with lateral margins much straighter than in S. tenuis , and in the nature of caudal rami seta IV, being unusually long and dilated (seta V was broken off in the specimen). With regard to the form of the genital double-somite, the species resembled O. tregoubovi , but the ornamentation of the maxillipedal basis (spatulated setules) and the armature of the antenna (seta IV absent) clearly distinguished it from this species and identified it as Spinoncaea . It may probably represent an as yet undescribed new species. In the Strait of Bab al Mandab (southern Red Sea) another morph of S. tenuis was found ( Spinoncaea sp. B, Table 3 View Table 3 ), which was close to the typical form, but differed slightly in the form of the genital double somite, being more rounded, and in the size of caudal seta III, being larger. The specimens resembled the elongate form of S. ivlevi , but differed from this type in the form of caudal rami seta IV, which was dilated (as in S. tenuis ), not setiform as in S. ivlevi . A specimen similar to this type was also found in the central northern Arabian Sea, where it had been grouped together with an as yet unidentified species of the tregoubovi -group under Oncaea sp. D in the earlier quantitative studies ( Böttger-Schnack, 1996). Another 2 specimens showing intermediate characters were found in the eastern Mediterranean ( Table 3 View Table 3 ), but their potential identity with Spinoncaea sp. B still needs to be examined. Specimens of Spinoncaea sp. B may be regarded as intermediate morphs of S. tenuis and S. ivlevi or they may represent another as yet undescribed species of Spinoncaea . More and detailed taxonomic studies of the tenuis -complex are needed in order to unravel its taxonomic diversity on a global scale, which still is far from being understood.
Ecological notes. Spinoncaea tenuis is the species previously reported as Oncaea sp. K in the Red Sea ( Böttger-Schnack, 1992, 1995). During the earlier ecological investigations in the Red Sea main basin ( Böttger-Schnack, 1988, 1990a,b) it had not been separated from the very similar O. tregoubovi Shmeleva which co-occurs in the upper mesopelagic zone. In a subsequent re-evaluation of the earlier data, the abundances and vertical distribution patterns of both species were established separately ( Böttger-Schnack, 1992).
Similar to S. ivlevi and other small oncaeid species, the actual abundance of S. tenuis had been largely underestimated in the earlier ecological studies in the central Red Sea during autumn and winter (e.g. Böttger-Schnack, 1992), because a net of 0.1 mm mesh had been used, which does not even sample the adults quantitatively (Böttger, 1985). Subsequent zooplankton studies in the central Red Sea conducted with a finer net of 0.055 mm mesh size during summer yielded between 3 and 10 times more adults (Böttger-Schnack, 1995). Also, the proportion of males differed considerably between the two surveys: they contributed 40–50% of adult S. tenuis in the 0.055 mm nets (unpubl. data), but only 0–5.0% of the adults in the 0.1 mm nets ( Böttger-Schnack, 1992). Besides some potential seasonal variation in abundance and sex ratio, the results pointed out a much more efficient catch of S. tenuis by the smaller mesh nets, in particular with regard to the males, which are somewhat smaller and more slender than the females.
Geographical distribution. Spinoncaea tenuis is distributed throughout the Red Sea main basin and in the Gulf of Aden ( Böttger-Schnack, 1995; as Oncaea sp. K). It has recently been recorded also from the Gulf of Aqaba, the northernmost extension of the Red Sea (Böttger-Schnack et al., 2001). In the Strait of Bab al Mandab at the southern entrance of the Red Sea, a considerable proportion of the unidentified specimens of the ivlevi -group reported by Böttger-Schnack (1995; as Oncaea ivlevi /sp. K) were assigned to S. tenuis upon recent re-examination. However, the samples also contained the elongate form of S. ivlevi and S. humesi , both of which are very similar in general aspect. Thus, quantitative data can hardly be provided, due to the difficulties in species identification that arise when all three species of Spinoncaea and their form variants co-occur. The present preliminary results pointed to a greater abundance of S. tenuis in the Strait of Bab al Mandab than previously reported by Böttger-Schnack (1995) and the regional differences in number between the South and the North in the Red Sea have to be corrected accordingly.
Outside the Red Sea, S. tenuis was recorded from a wide range of localities, including Atlantic as well as Indo-Pacific regions ( Table 3 View Table 3 ). The small mesh net samples from the Equatorial Indian Ocean (Nishida material) examined during the present study contained only few oncaeids similar to S. tenuis , which were not investigated further. In the copepod material from the SE Indian Ocean, near-shore area of the NW Cape of Australia (McKinnon material), S. tenuis was absent, which might indicate an oceanic distribution of the species.
Vertical distribution and vertical migration. In the Red Sea, S. tenuis is mainly distributed in the lower epi- and upper mesopelagic zone between 150 and 600 m depth, with maximum abundances in the 200–350 m layer ( Böttger-Schnack, 1992, 1995, and unpubl. data). It usually exhibits an unimodal distribution in this depth layer, with little regional variation in oceanic sites (unpubl. data from summer 1987). Thus, the species tends to be vertically separated from its two congeners, S. ivlevi and S. humesi . No diurnal vertical shift of the population centre was observed ( Böttger-Schnack, 1992; females only). The vertical extension of males usually was narrower than that of the females. In the shallow southern Red Sea and the Strait of Bab al Mandab, female S. tenuis were concentrated in the near-bottom layer, at 150–175 m and 150–200 m, respectively, during summer. Males showed a greater vertical extension in the shallow southern Red Sea with constant concentrations between 40 and 175 m depth; they were rare in the Strait. However, due to the difficulties in the identification of the species of the ivlevi -group (see above) the distribution pattern in the Strait might not have been fully discerned.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |