
Colobaea punctata (Lundbeck), 1923
|
publication ID |
https://doi.org/10.11646/zootaxa.4840.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:56993BCA-1A3E-415E-A765-0D55AB3E7A97 |
|
DOI |
https://doi.org/10.5281/zenodo.4478732 |
|
persistent identifier |
https://treatment.plazi.org/id/7170D74F-6A1E-FFCA-FF4F-FE898FFCDA79 |
|
treatment provided by |
Plazi |
|
scientific name |
Colobaea punctata (Lundbeck), 1923 |
| status |
|
Colobaea punctata (Lundbeck), 1923 View in CoL
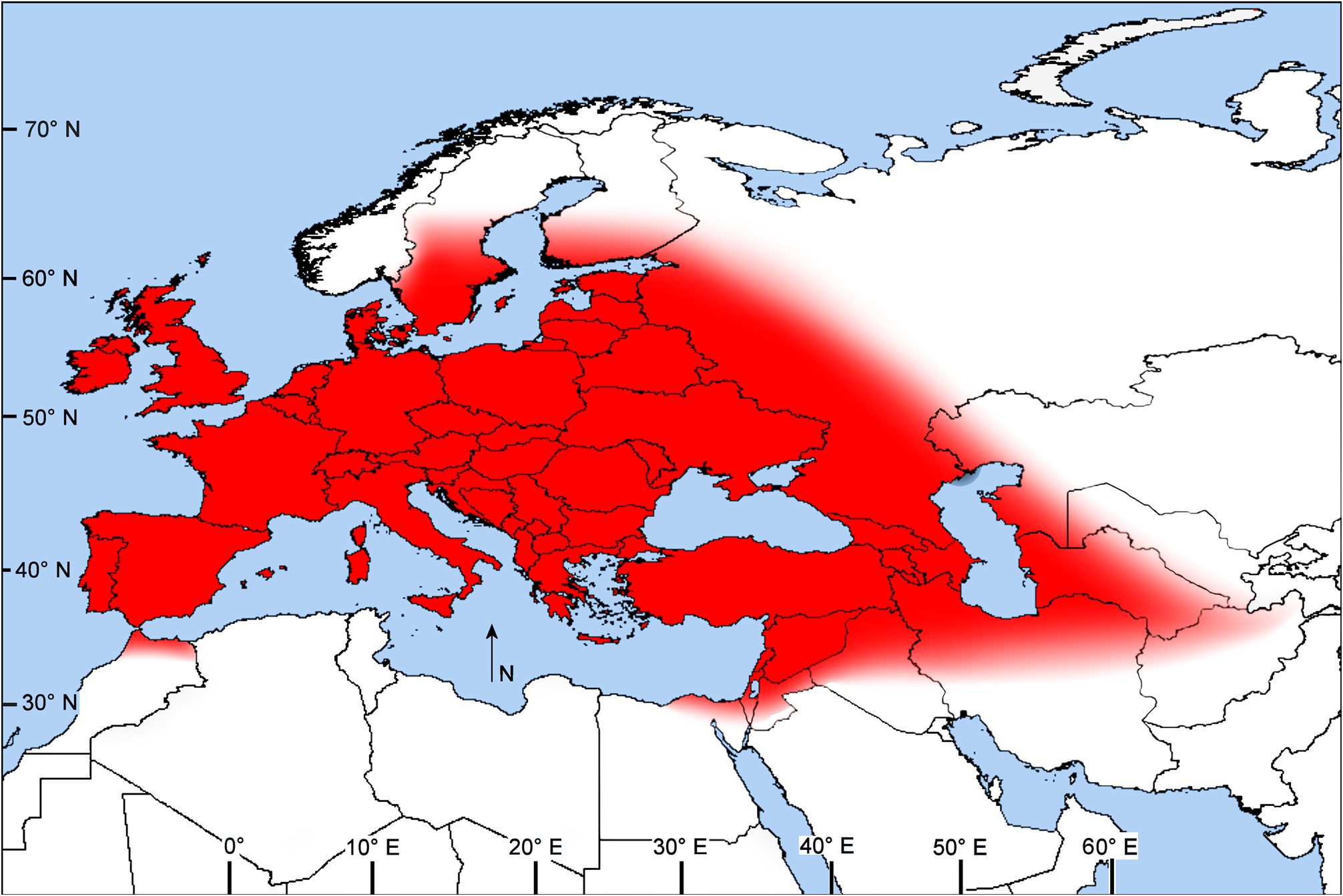
Map 15 View MAP 15
(BNN 6411; FT 6411, 6416, 6419–6420)
Adult flies of Colobaea punctata are very similar to those of C. pectoralis but differ noticeably in the extent of black pigmentation on the pleura and in characters of the male postabdomen. Adult flies of C. punctata have a discrete black spot below the anterior notopleural seta, whereas those of C. pectoralis have a black stripe extending along the upper margin of the mesopleuron to the base of the wing (see photograph at https://upload.wikimedia.org/wikipedia/commons/c/c7/ Colobaea _punctata.jpg). Rozkošný (1959) and Rivosecchi (1992) both provided a figure of an adult female.
Type locality. Rozkošný (1984b) noted, “The type-series [in MZLU] consists of 10 males and 10 females, but only four of them have locality labels (Donse and Damhusmosen in Denmark)” [bogs near Hillerød and Copenhagen, Zealand, Denmark]. Although C. punctata seems to have about the same northern limits (Jakobstad, Finland) as the closely related C. pectoralis , it extends much farther south, with a southeastern extension that is fairly continuous to Lake Ohrid, which lies between North Macedonia and eastern Albania. We have seen 1♂ of C. punctata from Turkey (Finike, Antalya); 1♀ from the northern Negev, Israel; 1♂ from Iran ( Mazanderan, 25 km west of Gorgan); 2♀ and 5♂ from Atvasar, Kazakhstan; and 1♂ from Bala Murghab Herat, northeastern Afghanistan ( Rozkošný & Knutson 2006). Rozkošný & Elberg (1991) recorded the species from “eastern Siberia” without providing a specific locality.
LVK reared C. punctata in southeastern Spain (Adra, near Almeria). The record from Almeria ( 37°N) on the Mediterranean Sea is about 1100 km south of the previously known southernmost locality (Montpellier, France; 43.5°N) in western Europe. Becker (1903) recorded C. pectoralis from Cairo, Egypt, but that was before C. punctata had been described; thus his record may have been based on a specimen of the latter species.
Laboratory rearings and observations are based on adult flies reared from puparia found in shells of P. planorbis collected between 15 February and 20 March 1964 at Adra, Spain.
The habitat distribution of the adult flies seems to be quite broad. Adult flies were collected along the partly shaded margin of a small lake (Wienerwald See) near Vienna, Austria (C.O. Berg in litt. to LVK). Coe (1958) recorded 1♂ “swept from vegetation by lake” at Lake Ohrid. Specimens in the ZMUC were collected in bogs in Zealand, Denmark, and at a millrace pond in Jutland, Denmark. Lundbeck (1923) found puparia in shells of several freshwater snails floating in spring flood refuse in Danish ponds and bogs. ADB and LVK discovered puparia in many shells of P. planorbis found floating among Carex , Juncus , and hygrophilous grasses emerging through 10–50 cm of water along the exposed shore of a small lake in the Albufera de Adra at Adra, Spain.
Adult flies mated within 24 hours after emergence from puparia and continued to mate from 9 March until 14 May, throughout the entire time during which the cultures of flies were maintained. The copulatory posture of the adult flies was found to be typical of that of most Sciomyzidae [described above under “ Colobaea bifasciella (Fallén), 1820 ”] and not unusual as in the closely related C. pectoralis . Even very gravid females were observed to copulate for long periods of time.
Females that emerged between 29 February and 18 April first oviposited as early as four days after emergence and at the latest within 17 days after emergence. They laid eggs between 14 March and 26 May on damp or wet vegetation or cotton in the breeding jars but laid no eggs on living P. planorbis that were also kept in the jars. Eggs hatched between 16 March and 30 May after incubation periods of 2–4 days at 19–21°C.
Of the five reared species of Colobaea , C. punctata appears to have the broadest food range. Lundbeck (1923) wrote, “This species I have bred from several snails, most often from young specimens of Planorbis planorbis L., and further from P. [ Planorbis ] albus Müll. and from youngs ( sic) of P. [ Planorbarius ] corneus L. and also from Lymnaea ovata Drap. var. peregra [ L. ovata = Radix balthica (L.)].” In contrast to Lundberg’s experience, all puparia found at Adra were in shells of P. planorbis . Small individuals of other species of aquatic snails [ Galba truncatula , Physella acuta (Draparnaud) , and R. peregra ] were abundant at Adra, but no puparia of C. punctata were found in shells of those species. During laboratory rearings, however, many C. punctata larvae killed, consumed, and developed to pupariation solely on those species of snails. Larvae also killed and consumed A. vortex , B. contortus , P. planorbis , Segmentina nitida (O.F. Müller) , and S. palustris . A few larvae attacked but did not kill small specimens of Succinea putris L. from Denmark. They did not attack “ Phystra myosotis ” [= Myosotella myosotis (Draparnaud) ] from Valencia, Spain, nor did any first-instar larvae initially feed on dead snails.
Newly hatched larvae crawled about actively and readily attacked snails. The methods of attack and feeding were similar for all species of snails consumed, but most of the detailed observations were made with small (about 5-mm-diam.) P. planorbis and small (about 4 mm long) P. acuta . Each snail observed during the rearings was attacked by a single first-instar larva; in crowded rearing containers, as many as 15 first-instar larvae would attack a single snail. It appeared that the young larvae fed on mucus, hemolymph, or extrapallial fluid rather than on snail tissues. An hour or so after the initial contact, depending upon the size of the host and the number of larvae that attacked it, the snail gradually began to retreat into its shell. The larvae followed, feeding at the edges of the mantle and foot. By the second day of feeding, the quiescent snail had withdrawn completely into its shell, and the larvae had penetrated between the mantle and shell until only their posterior spiracles remained exposed. A 5-mm-diam. P. planorbis that had been attacked by 10 first-instar larvae lived for three days after the initial attack. A 3-mm-long P. acuta lived for five days after being attacked by three first-instar larvae. Larvae continued to feed in the decaying tissues even after the tissues had become black and putrid until most of the contents of the shells had been consumed. Larval feeding appeared to be unaffected by the large populations of nematodes that often developed in the putrid snails. Older larvae attacked vigorously, immediately penetrated between the shell and the mantle in the acute corner of the aperture of P. acuta , and killed the snail within a few hours. The larvae killed and partially consumed a second and even a third snail if the first snail had not provided sufficient nourishment for completion of larval development. The total duration of larval life for 19 individuals was 9–16 days (average 10.3 days) at 19–22°C.
Larvae of C. punctata were reared to pupariation on P. planorbis , P. acuta , and S. palustris . Most larvae pupariated within shells even if copious liquefied tissue remained in the aperture. Only one puparium was formed in each shell. A few larvae pupariated on or under damp cotton or moss in the rearing containers. Shells of P. planorbis collected at Adra that contained puparia were 3.3–7.2 mm in diameter. Pupariating larvae excreted a small quantity of septum material, but on no occasion was the material formed into even a rudimentary septum as with C. americana and some species of Pherbellia . In the laboratory, 24 puparia were formed between 28 March and 25 April 1964, from which 19 adult flies ( 10♀, 9♂) emerged between 12 April and 12 May after puparial periods of 15–20 days (average 17.4 days) at 19–22°C.
Colobaea punctata apparently overwinters in the puparium. Lundbeck (1923) reared adult flies from puparia collected in the spring in Denmark. From 152 puparia that LVK collected between 15 February and 20 March 1964 in southern Spain in shells of P. planorbis , 99 adult flies ( 46♀, 53♂) emerged between 29 February and 17 June. Females emerged between 9 March and 18 April, 5–45 days (average 16.3 days) after the puparia had been collected, and males emerged between 29 February and 17 July, 6–89 days (average 18.2 days) after the puparia had been collected. LVK encountered neither adult flies nor other immature stages at Adra even when great numbers of puparia were present .
Of 40 laboratory-reared adult flies, 19♂ lived 4–26 days (average 14.0 days) and 21♀ lived 7–31 days (average 15.0 days).
Data from capture records of adult flies extend from 8 April (Finike, Antalya, Turkey) to 26 September (Berlin area; Kassebeer 2001). In his study of the flight periods and voltinism of the Sciomyzidae as based on museum material, Soós (1958) recorded 18 specimens of C. punctata collected between 15 July and 17 September. He concluded that C. punctata is a species “die sicher nur einer Generation besitzt” [“which surely has only one generation”].
Lundbeck (1923) wrote that he “reared a Cryptine and a Chalcidid from Ctenulus punctatus , in all cases a single parasite from each pupa.” Sixteen of 152 puparia collected at Adra produced a single parasitic wasp ( Hymenoptera : Chalcididae ). The wasps emerged between 19 March and 26 May, 15–67 days after the C. punctata puparia harboring them had been collected.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SuperOrder |
Heterobranchia |
|
Order |
|
|
SuperFamily |
Succineoidea |
|
Family |
|
|
Genus |