
Dracoderes nidhug, Thomsen, Vibe G., Rho, Hyun Soo, Kim, Dongsung & Sørensen, Martin V., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3682.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:1B811565-C78A-4CF2-9B89-723A20524280 |
|
DOI |
https://doi.org/10.5281/zenodo.5620879 |
|
persistent identifier |
https://treatment.plazi.org/id/6F11A301-1244-2231-FF53-8EB8F488FBC2 |
|
treatment provided by |
Plazi |
|
scientific name |
Dracoderes nidhug |
| status |
sp. nov. |
Dracoderes nidhug new species
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 , Tables 1–2 View TABLE 1 View TABLE 2 )
Diagnosis. Dracoderes with dorsal spines on segments 3 to 9, alternatingly offset to lateral sides on segments 3 to 8, subdorsal tubules on segment 2, lateroventral tubules on segments 2, 5 and 10, and lateral accessory tubules on segment 8. Tergal extensions of terminal segment with thin, flexible tip. Males with two large penile spines and a minute, lateroventral fringed spine.
Type material. Holotype, adult female, collected on November 11, 2011 in the East Sea, off Uljin, locality MAP37: 37°01ʹ26ʹ N, 129°30ʹ55ʹ E, from mud at 160 m depth; mounted in Fluoromount G®, deposited at the NHMD, under accession number: ZMUC KIN- 649. Allotype, adult male, same collecting data as holotype, mounted in Fluoromount G®, deposited at NHMD under accession number: ZMUC KIN- 650. Paratypes, four adult females and three adult males, collected on same date and locality as holotype, mounted in Fluoromount G®, deposited at NHMD under accession numbers: ZMUC KIN- 651 to KIN- 657. Additional material includes nine specimens mounted for SEM, stored in the personal collection of the last author.
Etymology. The species is named after the dragon “ Nidhug ”, known from Nordic mythology. At the End of Days, Nidhug will bite and cut the roots of Ash Yggdrasil – the Tree of Life.
Description. Adult specimens with of a head, a neck and eleven trunk segments ( Figs 1 View FIGURE 1 A–B, 2A, 3A, C).
Measurements and dimensions are given in Table 1 View TABLE 1 , and a summary of sensory spot, spine, tubule and glandular cell outlet positions is provided in Table 2 View TABLE 2 .
The head consists of a retractable mouth cone and an introvert. Inner armature of mouth cone with pharyngeal crown, one ring of helioscalids, and two rings of inner oral styles. The exact number of helioscalids and inner oral styles could not be determined. Outer mouth cone armature with nine outer oral styles that alternate in size between large well-developed ones in positions anterior to odd numbered introvert sections, and much smaller ones anterior to even numbered sections ( Fig. 3 View FIGURE 3 B). An outer oral style is missing anterior to introvert section 6. Large styles are composed of two joined units; distal unit terminates into hook-like structure ( Fig. 3 View FIGURE 3 B). Small styles are extremely under-developed, and resemble the soft inner oral styles. The introvert carries rings of scalids, but even though all specimens mounted for SEM had their introverts extended, scalids always tended to be collapsed and smacked together, so further examination was not possible.
Neck with nine placids, with a joint between the placids and segment 1 ( Figs 1 View FIGURE 1 A–B, 2B). The widest placids are the midventral and midlateral ones, ca. 19 µm at base, whereas the widths of the remaining ones vary between 16 µm and 18 µm. The 5 most ventral placids are situated closely together, whereas the midlateral, laterodorsal and subdorsal placids have cuticular foldings occupying the space between them ( Fig. 1 View FIGURE 1 A).
The first segment forms a closed cuticular ring, whereas the following 10 are composed of a tergal and two sternal plates ( Figs 2 View FIGURE 2 A, 3C). Junctions between tergal and sternal plates are easily observed with LM (e.g. Fig. 2 View FIGURE 2 C), but in SEM only the midsternal junction is marked by a visible line, whereas a continuous outer cuticle is present in the position of the tergosternal junctions, indicating that the plate junctions are intracuticular. Maximum sternal width is reached at segment 5, and the sternal widths taper gradually from this segment and towards the posterior end. The overall trunk outline appears cigar-shaped, rather than rectangular ( Fig. 2 View FIGURE 2 A).
Segment 1 without spines or hair. Sensory spots are present in subdorsal, sublateral and ventrolateral positions near the IJ-line, and more medially on the segment in laterodorsal, midlateral and ventromedial positions ( Figs 1 View FIGURE 1 A– B, 3D). Sensory spots on this and all following segments are rather small and rounded, with cuticular papillae. The cuticle appears smooth, except on the ventral side where two indistinct markings form a line with presumably thinner cuticle ( Figs 2 View FIGURE 2 B, 3D). The lines run from ventrolateral positions at the anterior segment margin and extend posteriorly and obliquely to a more ventromedial position. At the level of the IJ-line, the folds bend towards each other, and meet midventrally. The folds are extremely weak in LM (but see Fig. 2 View FIGURE 2 C), but were visible in all specimens prepared for SEM. The posterior segment margin is straight along the dorsal and lateral sides, but forms a broad, midventrally pointed extension ( Figs 2 View FIGURE 2 B, 3D). A pair of small indentations in the posterior margin is present in line with the tergosternal junctions of the following segment. Except at the indentations, the segment edge forms a finely serrated fringe around the segment ( Fig. 3 View FIGURE 3 D, E–F).
Segment 2 without spines, but with small tubules in subdorsal and lateroventral positions ( Figs 1 View FIGURE 1 A–B, 2C–D, 3E–F). Sensory spots are present as twin pairs in laterodorsal position, and single pairs in ventrolateral and ventromedial positions. The ventrolateral pair is placed towards the anterior end of the segment, whereas the rest are located medially on the segment. Glandular cells are present in paraventral positions on this and all following segments, down to segment 10. Indentations are present in the posterior segment margin in same position as on the preceding segments, but on these and the following segments, the indentations are not as deep as those in segment 1. Cuticular hairs are arranged in two rows that extend from subdorsal to ventromedial positions, with the shortest hairs in the anterior row ( Figs 1 View FIGURE 1 A, 3E). The rows are interrupted midlaterally by hairless spaces. Anterior part of segment, near overlapping free flap from preceding segment, with secondary fringe resembling a curtain with fine, posterior serration. Posterior segment margin serrated, with alternatingly larger and smaller teeth.
Segment 3 with a single dorsal spine, slightly displaced to the left or right, into a more lateral position ( Figs 1 View FIGURE 1 A, 2D, 3A); dorsal spines on this and following six segments are thin and acicular, composed of a basal sheath, and an acicular end piece with finely serrated margins. Trunk cuticle around the attachment points of dorsal spines with conspicuous subcuticular lateral extensions that are best observed with LM ( Fig. 2 View FIGURE 2 D). The proximal part of the spine attaches to the trunk cuticle through a spherical, condyle-like structure, suggesting a ball-and-socket articulation. Spine with perispinal paradorsal sensory spots ( Fig. 1 View FIGURE 1 A). Additional sensory spots are located in laterodorsal, sublateral, ventrolateral and ventromedial positions. Hairs are located as on the preceding segment, but with an additional short row placed above the other two from subdorsal position and across the hairless gap. Glandular cell outlets, secondary fringe and posterior segment margin as on preceding segment.
Segment 4 with single dorsal spine, slightly displaced to a more lateral position, opposite to the side of the dorsal spine on the preceding segment. Hairs are arranged in three rows, all interrupted by the same hairless areas as on segment 2; however with a small laterodorsal patch of hair in each row. Sensory spots, glandular cell outlets, secondary fringe and posterior segment margin as on preceding segment.
Segment 5 with single dorsal spine, slightly displaced to a more lateral position, opposite to the side of the dorsal spine on the preceding segment. A pair of short tubules is present in the lateroventral positions ( Figs 1 View FIGURE 1 B, 2F). Sensory spots, glandular cells, cuticular hairs, secondary fringe and posterior segment margin as on preceding segment.
Segment 6 with single dorsal spine, slightly displaced to a more lateral position, opposite to the side of the dorsal spine on the preceding segment. A pair of acicular spines present in the lateroventral positions ( Fig. 1 View FIGURE 1 B); lateroventral spines are composed as the dorsal ones, with proximal ball, basal sheath and acicular end piece, but appear somewhat shorter and stouter. Trunk cuticle strongly reinforced around the attachment point of the lateroventral spines, creating conspicuous subcuticular thickenings and ventrally directed extensions (see similar from segment 8 on Fig. 2 View FIGURE 2 E). Similar extensions are present around the lateroventral spines on all following segments. Cuticular hairs appear as on the preceding segment, but are sparser and in only two rows. Sensory spots, glandular cells outlets, secondary fringe and posterior segment margin as on preceding segment.
Segment 7 is identical with preceding segment, except for the dorsal spine that is laterally displaced to the opposite side, the ventromedial sensory spots that are absent and cuticular hairs being reduced to a patch of two rows placed laterodorsally, and a few single hairs subdorsally.
Segment 8 identical with segment 6, except for the presence of short lateral accessory tubules ( Figs 1 View FIGURE 1 B, 2E, 3G), and the cuticular hairs now only being present as a few subdorsal ones.
Segment 9 with dorsal and lateroventral spines ( Fig. 3 View FIGURE 3 G). The dorsal spine is located in a perfect middorsal position. Nephridial pores present in lateral accessory positions ( Figs 1 View FIGURE 1 B, 2E). The segment is hairless. Sensory spots, glandular cells outlets, secondary fringe and posterior segment margin as on preceding segment.
Segment 10 without spines and cuticular hairs, but with pair of lateroventral tubules ( Fig. 2 View FIGURE 2 B–C). Sensory spots present in subdorsal, laterodorsal, sublateral (males only) and ventrolateral positions. Glandular cells outlets, secondary fringe and posterior segment margin as on preceding segment.
Segment 11 with lateral terminal spines. Males with two flexible and long (60–70 µm) penile spines emerging laterally from the intersegmental joint between segment 10 and 11 ( Figs 1 View FIGURE 1 C, 2G–H, 3H–I). In most specimens the two penile spines tended to be stuck together, appearing as one spine with a grove, but one specimen clearly showed that the spines are separated (Fig, 3I). Males furthermore with a pair of short (14 µm), lateroventral flexible spines, with tiny fringes arranged around them as rings ( Figs. 1 View FIGURE 1 C–2H, 3H–I). Females do not show any outer, sexual dimorphic characters ( Fig. 1 View FIGURE 1 A–B). Two pairs of subdorsal sensory spots present, one on the anterior part of segment, and the second one towards the tips of the tergal extensions. Sensory spots furthermore present in ventrolateral positions near the lateral, posterior corners of the sternal plates ( Fig. 3 View FIGURE 3 H). Glandular cells and cuticular hairs are not present. Secondary fringe present dorsally, on some specimens stretching nearly halfway down the segment. Posterior margin of tergal plate bulges before extending into triangular tergal extensions, extending well beyond the truncate sternal plates ( Figs 1 View FIGURE 1 A–C, 2H, 3H). Dorsal side of tergal extensions smooth ventral side densely covered with short, papillar hairs ( Figs1 View FIGURE 1 C, 3H). Each tergal extension has a flexible, spinous elongation extending from the tip ( Fig. 2 View FIGURE 2 H).
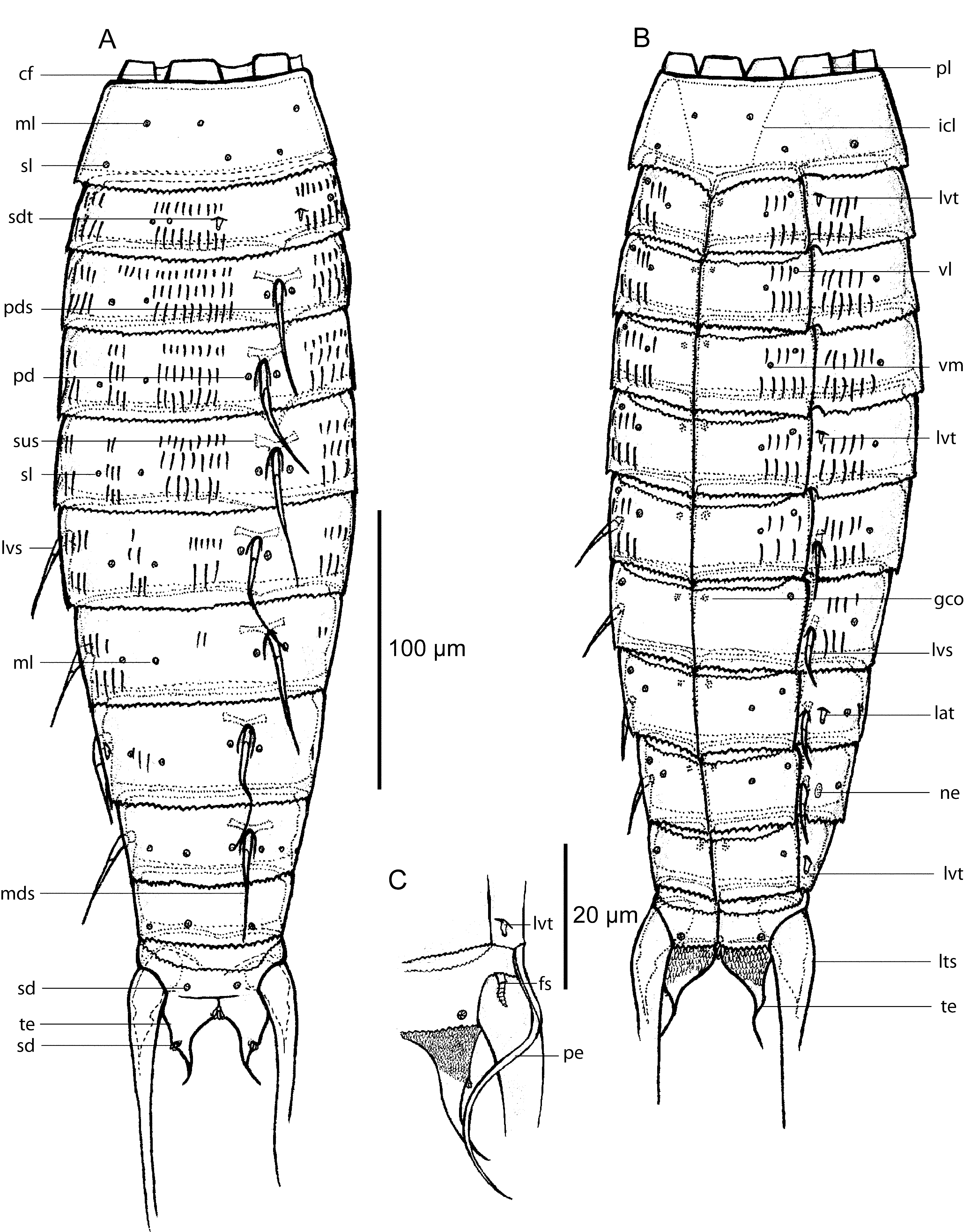

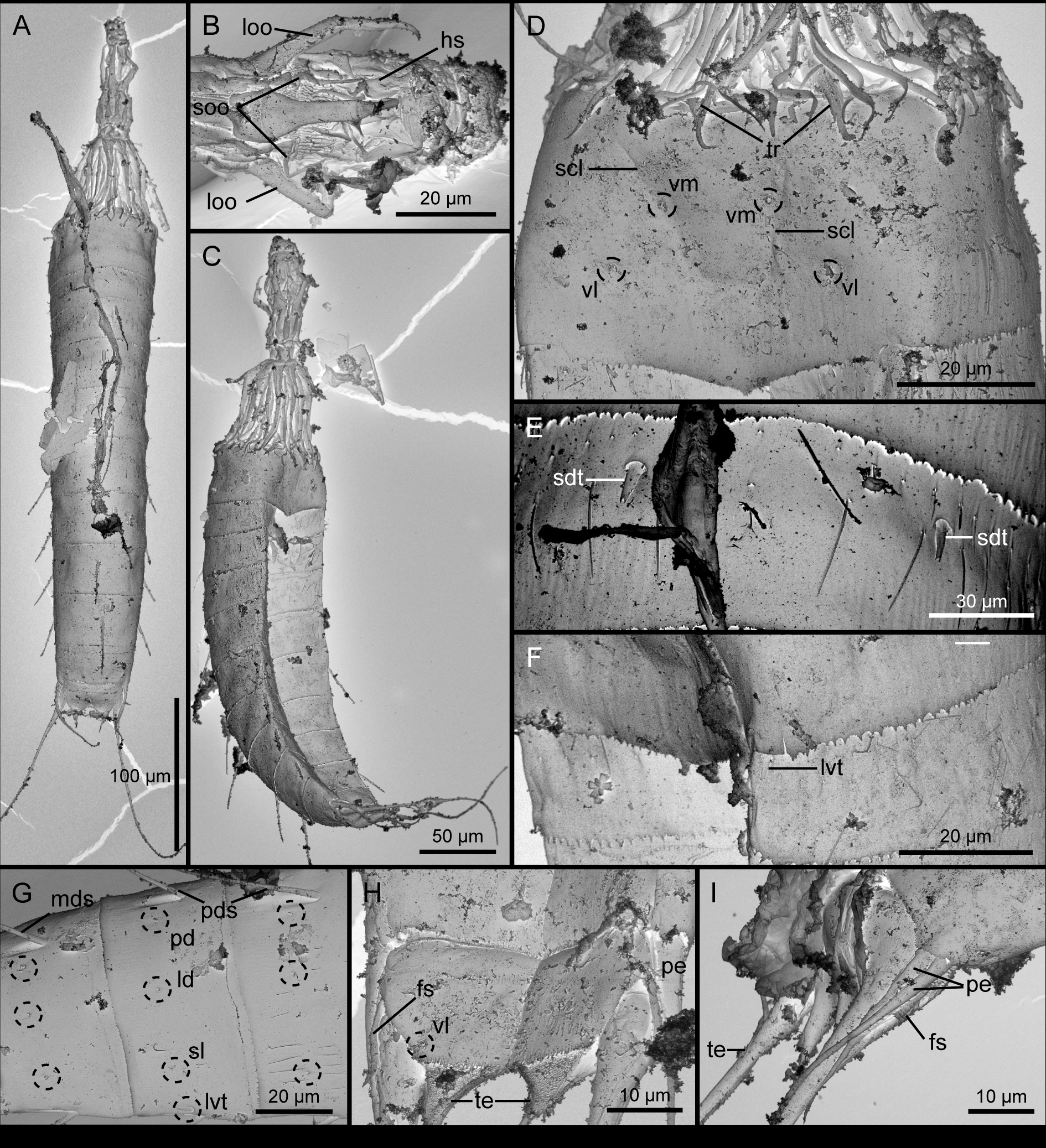
TABLE 1. Measurements of two adult specimens of Dracoderes nidhug nov. sp. from the East Sea, Korea, including number of measured specimens (n) and standard deviation (S. D.). Abbreviations: (ac): acicular spine; LTS: lateral terminal spine; LV: lateroventral; MD: middorsal spine; MSW- 5: Maximum sternal width, measured on segment 5 in this species; PD: dorsal spine displaced to paradorsal position; S: segment length; SW- 10, standard width, always measured on segment 10; TL: trunk length.
| Character | n Range | Mean | S.D. |
|---|---|---|---|
| TL | 9 374–427 µm | 403 µm | 15,85 µm |
| MSW-5 | 9 87–95 µm | 90 µm | 3,14 µm |
| MSW-5/TL | 9 21,0–23,3 % | 22.3 % | 0,68 % |
| SW-10 | 9 59–68 µm | 65 µm | 3,21 µm |
| SW-10/TL | 9 14,9–17,0 % | 16.0 % | 0,84 % |
| S1 | 9 43–48 µm | 46 µm | 1,58 µm |
| S2 | 9 36–40 µm | 38 µm | 1,32 µm |
| S3 | 9 37–41 µm | 38 µm | 1,22 µm |
| S4 | 9 39–44 µm | 41 µm | 1,88 µm |
| S5 | 9 39–47 µm | 43 µm | 2,33 µm |
| S6 | 9 40–46 µm | 43 µm | 2,09 µm |
| S7 | 9 39–45 µm | 42 µm | 2,24 µm |
| S8 | 9 36–43 µm | 41 µm | 2,30 µm |
| S9 | 9 36–43 µm | 40 µm | 2,24 µm |
| S10 | 9 29–33 µm | 32 µm | 1,33 µm |
| S11 | 9 39–47 µm | 43 µm | 2,42 µm |
| MD 3 (ac) | 9 43–57 µm | 50 µm | 4,85 µm |
| PD 4 (ac) | 9 40–62 µm | 55 µm | 7,64 µm |
| PD 5 (ac) | 9 40–70 µm | 58 µm | 9,55 µm |
| PD 6 (ac) | 8 55–63 µm | 59 µm | 2,75 µm |
| PD 7 (ac) | 9 44–57 µm | 51 µm | 4,40 µm |
| PD 8 (ac) MD 9 (ac) | 8 35–47 µm 9 48–57 µm | 43 µm 53 µm | 4,34 µm 3,35 µm |
| LV 6 (ac) | 9 30–37 µm | 34 µm | 2,12 µm |
| LV 7 (ac) | 9 30–36 µm | 34 µm | 2,35 µm |
| LV 8 (ac) | 9 27–33 µm | 31 µm | 1,80 µm |
| LV9 (ac) LTS | 9 26–31 µm 8 200–229 µm | 28 µm 215 µm | 2,19 µm 11,41 µm |
TABLE 2. Summary of nature and location of sensory spots, glandular cell outlets, spines and tubules arranged by series in Dracoderes nidhug nov. sp. Abbreviations: LA: lateral accessory; LD: laterodorsal; LV: lateroventral; MD: middorsal; ML: midlateral; PD: paradorsal; PV: paraventral; SD: subdorsal; SL: sublateral; VL: ventrolateral; VM: ventromedial; ac: acicular spine; fs: fringed spine; gc, glandular cell outlet; lts: lateral terminal spine; (m): male condition of sexually dimorphic character; ne, nephridiopore; (pe): marks that the structure is perispinal and its exact position along the longitudinal series may have been obscured by eventual lateral spine displacement; ps, penile spine; ss, sensory spot; tu, tubule.
| Position Segment MD | PD | SD LD | ML LA SL | LV | VL | VM | PV |
|---|---|---|---|---|---|---|---|
| 1 | ss ss | ss ss | ss | ss | |||
| 2 | tu ss,ss | tu | ss | ss | gc | ||
| 3 ac | ss, | ss | ss | ss | ss | gc | |
| 4 | ac, ss (pe) | ss | ss | ss | ss | gc | |
| 5 | ac, ss (pe) | ss | ss | tu | ss | ss | gc |
| 6 | ac, ss (pe) | ss | ss | ac | ss | ss | gc |
| 7 | ac, ss (pe) | ss | ss | ac | ss | gc | |
| 8 | ac, ss (pe) | ss | tu ss | ac | ss | ss | gc |
| 9 ac | ss | ss | ne ss | ac | ss | ss | gc |
| 10 | ss ss | ss (m) | tu | ss | gc | ||
| 11 | ss,ss | ps,ps (m) lts | fs (m) | ss |
| ZMUC |
Zoological Museum, University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |