
Phractocephalus nassi, Lundberg & Aguilera, 2003
|
publication ID |
https://doi.org/ 10.1590/S1679-62252003000200004 |
|
persistent identifier |
https://treatment.plazi.org/id/677287A3-A229-FFE4-FEE1-568CFC78FB3F |
|
treatment provided by |
Carolina |
|
scientific name |
Phractocephalus nassi |
| status |
sp. nov. |
† Phractocephalus nassi , new species
Figs. 3 View Fig , 5 View Fig a-b, 5e-h, 5k-l
Phractocephalus hemiliopterus (misspelling) Lundberg et al. 1988:131-138 (description, phylogeny, biogeography); Lundberg 1997:80 (listed).
Phractocephalus Lundberg & Chernoff. 1992:10 View in CoL (listed). Phractocephalus hemioliopterus Lundberg 1998:59-62 View in CoL
(phylogeny, biogeography).
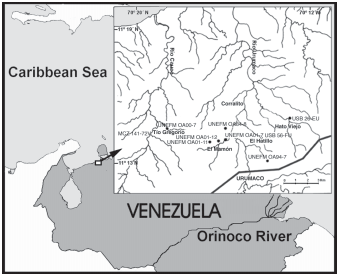
Holotype. MCN. USB OL-2142 , nearly complete, undistorted neurocranium plus most of the Weberian complex (broken into two pieces at the basioccipital). El Mamón oil field, 350 m north of oil well number 1, locality USB 56 View Materials - FU, 11º 15´N, 70º 13´W, Urumaco Formation , middle member, Falcón State, northwestern Venezuela, O. Linares, E. Pannier & R. Sánchez, 1987. GoogleMaps
Paratypes. All localities in Falcón State, northwestern Venezuela ( Fig. 2 View Fig ). MCN. USB 72-85 View Materials PB, nearly complete neurocranium, margins partly chipped or eroded (specimen described by Lundberg et al., 1988). Locality USB 26 View Materials - FU, 11º 15´00´´ N, 70º 13´00´´ W, Urumaco Formation, upper member. UNEFM-PF-0365, three dimensional Weberian complex and supraoccipital. Tío Gregorio, locality UNEFM OA00-7, 11º, 14´31´´ N; 70º 18´40´´ W, Urumaco Formation, upper member, O. Aguilera & R. Sánchez, 2000 GoogleMaps .
UNEFM-PF-0158, cranium in two large and several small parts; slightly crushed and flattened, most margins chipped or broken. Anterior moiety includes mesethmoid, lateral ethmoids, frontals and sphenotics. Ventrally includes vomer and parasphenoid. Smaller posterior part includes supraoccipital (anterior part to frontal and partial supraoccipital process with all margins chipped). Associated bones including nearly complete right hyomandibula, right quadrate, left opercle and left cleithrum. El Hatillo, locality UNEFM OA94-7, 11º, 13´03´´ N; 70º 14´06´´ W, Urumaco Formation, middle member, O. Aguilera & R. Sánchez, 1994.
UNEFM-PF-0363, right side, articulated, cleithrum, coracoid and pectoral spine. El Mamón, locality UNEFM OA01-11 View Materials , 1000m west of oil field, upper Miocene Urumaco Formation (middle member), Falcón State, northwestern Venezuela, O. Aguilera & J. Lundberg, 2001 .
UNEFM-PF-0311, partial left pectoral spine shaft (base missing). Quebrada El Mamón, locality UNEFM OA94-8 View Materials , 11º 14´27´´ N, 70º 16´00´´ W, Urumaco Formation, middle member, O. Aguilera & R. Sánchez, 1994 GoogleMaps .
Non-type specimens. UNEFM-PF-0366 skull fragment. Tío Gregorio, locality UNEFM OA00-7, 11º 14´31´´ N, 70º 18´40´´ W, Urumaco Formation, upper member, O. Aguilera & R. Sánchez, 2000 GoogleMaps .
UNEFM-PF-0364 supraoccipital fragment. El Mamón, 500 m west of oil field, locality UNEFM OA01-12, Urumaco Formation, middle member, O. Aguilera & R. Sánchez, 2001.
MCC-URU-2002-141, cranium in two large parts; slightly crushed and flattened, most margins chipped or broken. Anterior moiety includes mesethmoid, lateral ethmoids, frontals and sphenotics. Smaller posterior part includes supraoccipital (anterior part to frontal and partial supraoccipital process with all margins chipped). Tío Gregorio between Quebrada Tío Gregorio and Arcas fault, locality MCZ 141-72 V, Urumaco Formation, upper member, B. Patterson, 1972.
UNEFM-PF-0159, nearly complete right cleithrum. Tío Gregorio, locality UNEFM OA00-7, 11º, 14´31´´ N; 70º 18´40´´ W, Urumaco Formation, upper member, O. Aguilera & R. Sánchez, 2000 .
UNEFM-PF-0356, left side suspensorium, reassembled articulated bones including nearly complete hyomandibula, quadrate and part of metapterygoid. El Mamón, 1000 m northeast of oil field, locality UNEFM OA01-7, Urumaco Formation, middle member, O. Aguilera, J. Lundberg & R. Sánchez, 2001.
Locality, geological context and age. The sample area ( Fig. 2 View Fig ) includes the following named localities: El Mamón, Tío Gregorio, Corralito and El Hatillo, all north of the town of Urumaco, Falcón State, northwestern Venezuela. The geological units from which the specimens were obtained are the middle and upper members of the Urumaco Formation of Venezuela. These units have been recently summarized in the Léxico estratigráfico de Venezuela (1997).
The middle member of the Urumaco Formation consists of claystone and sand. The gray claystone is microfossiliferous, and the brown claystone contains vertebrate remains such as reptiles, mammals, marine and freshwater fish, and also coprolites and wood (Díaz de Gamero & Linares, 1989). The limestone changes from conchiferous sandstones to consolidated coquina limestone. In the lower half of this middle member, an abundant and diverse marine mollusk fauna exists in a sandy matrix. This matrix also contains elasmobranch teeth and teleostean otoliths. The paleoenvironments are interpreted as inner sub-littoral and coastal lagoon enviroments with riverine and estuarine influence ( Díaz de Gamero, 1996).
The upper member of the Urumaco Formation also comprises gray to brown often limey claystone with thin intercalated and locally conchiferous sandstones. The uppermost layer is referred to as the “capa de tortugas” because of its abundant remains of the turtle Bairdemys Gaffney & Wood. Several localities and levels have concentrations of vertebrate fossils. The vertebrate fauna includes marine, estuarine and freshwater fishes, terrestrial, freshwater and marine turtles and crocodilians, and terrestrial and aquatic/semiaquatic mammals ( Sánchez-Villagra et al., 2003). The paleoenvironments were tropical near shore marine to low coastal savannas with freshwater rivers.
A Late Miocene age for the Urumaco Formation is indicated by foraminiferans, and the mammals suggest a South American land mammal age between Chasicoan and Huayquian (Díaz de Gamero & Linares, 1989), or approximately 9 Ma (Marshall & Sempere, 1993).
Diagnosis. 1) Ornamentation of posterior half of frontals and anterior half of supraoccipital with both reticulating ridges and pits, and equally coarse, elongate, nearly parallel ridges and sulci ( Fig. 3a View Fig ).
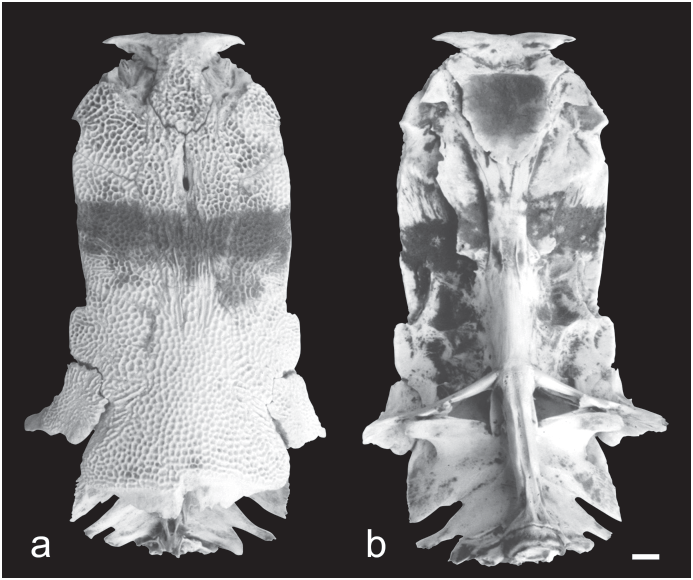
In modern P. hemioliopterus the corresponding central region of the skull roof is almost completely ornamented by reticulating ridges and pits, with few or no elongate, parallel ridges and sulci ( Fig. 4a View Fig ). In the Acre Phractocephalus the central region of the skull roof has many elongate ridges and sulci plus reticulating ridges and pits.
2) Mesethmoid bone very broad ( Table 1); dorsal surface extensively ornamented; anterior edge scarcely emarginate to gently rounded ( Fig. 3a View Fig ).
The mesethmoid bone of P. hemioliopterus is relatively narrow-necked, less extensively ornamented only centrally and posteriorly, and its anterior edge is emarginate or shallowly concave ( Fig. 4a View Fig ). The mesethmoid bone of the Acre Phractocephalus is similar to that of † P. nassi .
3) Lateral ethmoid not excessively projecting anteriorly over palatine condyle; its lateral margin convex and eliminating orbital notch on margin of skull roof ( Fig. 3a View Fig ).
The lateral ethmoid bone of P. hemioliopterus is also not excessively projecting anteriorly over the olfactory region or palatine condyle; but it is concave laterally, and has a well defined orbital sulcus on the margin of skull roof ( Fig. 4a View Fig ). In the Acre Phractocephalus the lateral ethmoid projects far anteriorly over the olfactory region and palatine condyle; it is convex laterally, eliminating the orbital sulcus on margin of skull roof.
4) Anterior cranial fontanelle absent or represented by a small pit in epiphyseal region between frontals ( Fig. 3a View Fig ).
Modern P. hemioliopterus have an open anterior cranial fontanelle within a prominent depression of epiphyseal region ( Fig. 4a View Fig ), whereas in the Acre Phractocephalus the fontanelle and depression of the epiphyseal region are lacking.
5) Supraoccipital process with rounded lateral and posterolateral margins, posterior margin concave medially; supraoccipital process long, in dorsal view completely covering modified anterior vertebrae of Weberian complex ( Fig. 3a View Fig ).
In modern P. hemioliopterus the supraoccipital process is short, in dorsal view not completely covering modified vertebrae of Weberian complex; it has angular lateral and posterolateral margins, and its posterior margin is irregular to concave medially and bearing a deep unornamented extension below anterior nuchal plate ( Fig. 4a View Fig ). In the Acre Phractocephalus the supraoccipital process is long, completely covering the Weberian complex; its process has angular corners and straight lateral margins, and its posterior margin concave is slightly convex and lacks an unornamented extension below anterior nuchal plate.
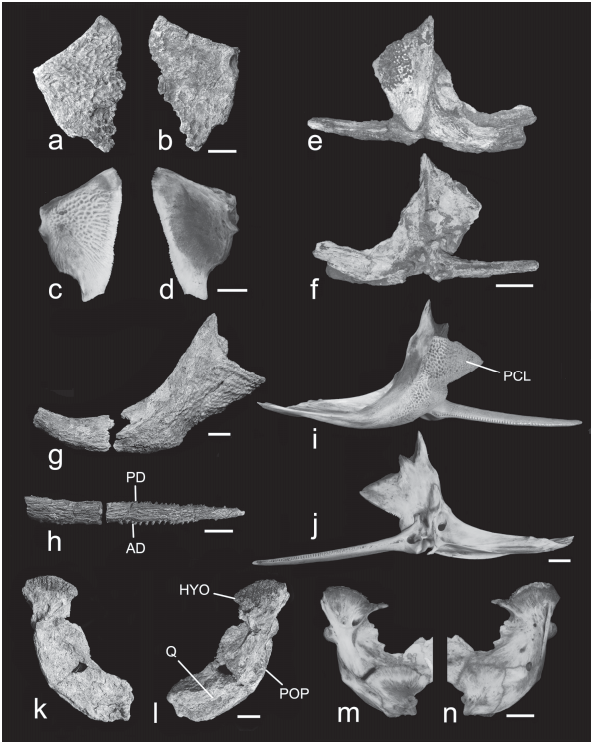
6) Opercle almost completely covered with reticulating ridges and pits ( Fig. 5a View Fig ).
The opercle P. hemioliopterus is incompletely covered with reticulating ridges and pits ( Fig. 5c View Fig ). The opercle is thus far unknown in the Acre Phractocephalus .
7) Cleithrum with coarse ornamentation extending along ventral edge forming outward bulge or “shoulder” lateral to spine articulation; bulge of “shoulder” terminating posteriorly, in line with termination of articular socket of pectoral spine ( Figs. 5 View Fig e-g).
The cleithrum of P. hemioliopterus lacks coarse ornamentation along ventral edge of “shoulder,” and the lateral bulge of “shoulder” terminates posterior to the level of articular socket of pectoral spine ( Figs. 5i, j View Fig ). No cleithrum of the Acre Phractocephalus has been found.
8) Pectoral spine ornamentation dominated by coarse subparallel ridges and sulci, and relatively few reticulating ridges and pits ( Fig. 5h View Fig ).
The pectoral spine ornamentation of P. hemioliopterus is dominated by reticulating ridges and pits ( Fig. 5i View Fig ). The pectoral spine of the Acre Phractocephalus is yet unknown.
Etymology. The Urumaco species of Phractocephalus is named for Dr. Pedro Nass in recognition of his contributions to the systematics and morphology of pimelodid catfishes of South America.
Description. See Lundberg et al. (1988) for description of neurocranium (MCN.USB 72-85 PB) and additional measurements. The following information is provided by the holotype and recently obtained specimens.
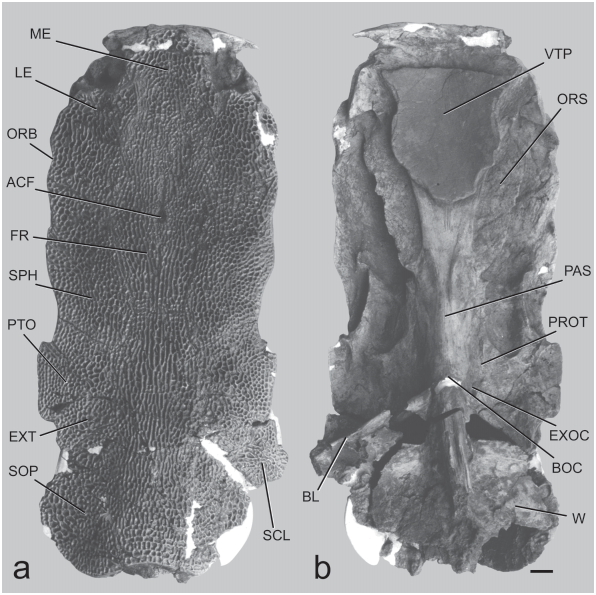
Neurocranium, dorsal aspect ( Fig. 3a View Fig ). Outline form of skull approximately rectangular, width across lateral ethmoids equal to width across sphenotics, and these widths slightly less than maximum width across pterotic wings; width across pterotics contained 2.25 times in (45% of) dorsal skull length from mesethmoid tip to posterior edge of supraoccipital process. Surface mostly flat but margins curved slightly downward and dorsal midline increasingly arched posteriorly. All bones ornamented with coarse reticulating ridges and circular pits and some longitudinally elongate ridges and grooves, except for olfactory capsules and extremities of mesethmoid cornua.
Mesethmoid broad, mushroom-shaped, anterior margin scarcely emarginate (entire in MCN.USB OL-2142), otherwise gently rounded; cornua stout, pointed, little recurved and reaching laterally to a parasagittal line projected before palatine condyle. Mesethmoid without traces of anterior cranial fontanelle. Lateral ethmoid projecting anteriorly into olfactory capsule region; orbital margin short, gently concave, antorbital process a slight curved bulge; contact with sphenotic a long, nearly straight suture running obliquely from skull roof margin behind orbit to frontal near anterior cranial fontanelle. Olfactory capsule floored by mesethmoid and lateral ethmoid. Nasal and infraorbitals unknown.
Frontal flat to slightly concave, confined to center of skull roof, laterally suturing to and excluded from skull-roof margin by lateral ethmoid and sphenotic. Anterior cranial fontanelle present as reduced circular foramen in a midline pit at transverse level of sphenotic-lateral ethmoid-frontal junction (fontanelle obsolete in MCN.USB OL-2142). Frontal ornamented mostly with reticulating ridges and circular pits, except near midline posterior to cranial fontanelle about 5 - 6 parallel longitudinally oriented ridges and grooves.
Sphenotic very large, twice the width of adjacent frontal, approaching a parallelogram in form, broadly sutured to lateral ethmoid, frontal, supraoccipital and pterotic; free lateral margin with a gently convex bulge but lacking prominent postorbital process. Pterotic with angular wing projecting from posterolateral corner; broadly contacting sphenotic, supraoccipital, extrascapula and supracleithrum, the last joint horizontally elongate suggesting limited mobility to dorsoventral axis.
Supraoccipital unusually large, flask-shaped; contacting frontals, sphenotics, extrascapulars, supracleithra and anterior nuchal plate (the last by inference from structure of posterior process and similarity to congeneric species). Expanded part of supraoccipital posterior process is posterior to occipital wall and articulations with upper shoulder girdle elements; process long, covering Weberian complex. Process with broadly rounded lateral and posterolateral margins, ornamented part posterior margin emarginate to truncate across midline, plus a projecting unornamented shelf that would form a lap joint with anterior nuchal plate. Ornamentation of supraoccipital mostly irregular but ridges and pits loosely arranged in 5 or 6 outer concentric rows. Dorsal surface flat behind frontals, then becoming convexly arched along midline to posterior margin, laterally sloping downward concavely to margins of posterior process; cross-sectional shape depressed “bell shaped.”
Extrascapula an ovoid plate isolated from skull margin by surrounding pterotic, supracleithrum and supraoccipital. Supracleithrum preserved in holotype on right side where broken posterolaterally; expanded as roughly lozenge-shaped plate, ornamented as the skull roof; horizontally elongate, weak sutural joint with pterotic and extrascapula; arthrosis style with supraoccipital indeterminate.
Neurocranium, ventral aspect ( Fig. 3b View Fig ). Mesethmoid visible ventrally only as flattened, narrow band, anterior to vomerine tooth patch, to which premaxillae (not preserved) articulate; mesethmoid cornua not downwardly deflected. Vomer dominated by massive median tooth plate, approximately pentagonal form, surface concave upward and completely covered by minute pediculate tooth-attachment bases (no teeth preserved). Vomer expanded antrolaterally in front of tooth plate to suture with mesethmoid and lateral ethmoids; tapering posterior limb deeply sutured with parasphenoid.
Palatine condyle projecting prominently from lateral ethmoid, very long and in horizontal plane, convex anteriorly, nearly straight laterally, abruptly truncate posteriorly. Prominent, thin, horizontal “orbital shelf” arises on ventral surface of lateral ethmoid just posterior to palatine condyle and lateral to vomer. “Orbital shelf” continuing onto orbitosphenoid, parasphenoid and prootic to terminate ventral to trigeminofacial foramen; along lateral ethmoid and orbitosphenoid “orbital shelf” is the surface of origin for adductor arcus palatini muscle. Orbitosphenoid dominated by “orbital shelves,” widest anteriorly, about 75% of skull width across lateral ethmoids, narrowing posteriorly to about 45% of skull width across sphenotics at level of hyomandibular facet. Foramina of orbitosphenoid deeply situated and indeterminate. Parasphenoid median stem broad, center of parasphenoid marked by medially converging pair of low ridges and elongate roughened surfaces (associated with contact of anterior branchial arches); parasphenoid weakly sutured to prootics, deeply sutured to basioccipital, indeterminate contact with pterosphenoid.
Anterior half of ventral surface of sphenotic planar and bare; posterior half of sphenotic sharply elevated laterally, forming anterior two-thirds of elongate, horizontal, troughlike hyomandibular facet that runs mediolaterally to sphenoticpterotic suture near skull roof margin, then along edge of pterotic. Hyomandibular facet on pterotic followed at rightangles by another articular facet, short, flat-faced and ventrally buttressed, for posterodorsal corner of hyomandibula. Neither pterosphenoid nor prootic participate in hyomandibular facet or other articulation with hyomandibula.
Medioventrally adjacent to hyomandibular facet, sphenotic, pterotic and prootic together form roof of ovoid, shallow subotic fossa.
Prootic side wall flat dorsal to contact with parasphenoid; position of matrix-filled trigeminofacial foramen indicated by semicircular notch in anterior border of prootic and sharp edged terminus of “orbital shelf” ventral to foramen. Pterosphenoid largely obscured by matrix but no evidence that it is more extensive than in modern species where it is restricted as a small quadrangular element between the prootic, sphenotic and orbitosphenoid.
Basioccipital weakly sutured to exoccipital and prootic; vertebral-like joint with first centrum. Articulation site on basioccipital for ossified Baudelot’s ligament elevated and rugose. Exoccipital weakly sutures with basioccipital, prootic, pterotic and epioccipital; contributing small dorsal process to cranial articulation with Baudelot’s ligament; vagal foramen large, circular, ventrally directed, centered on a vertical through the anterior edge of basioccipital-Baudelot’s ligament joint. Baudelot’s ligament of supracleithrum ossified and heavy; round in section medially near contact with basioccipital and exoccipital. Exoccipital and epioccipital forming sharp posterolateral corner of braincase that vertically buttresses expanded cranial articulation of pteroticsupracleithrum. Anterolateral face of epioccipital concave and weakly sutured to pterotic. Posterior end of pterotic wing produced and expanded ventral to cranial articulation of supracleithrum. Ventral side of supraoccipital posterior process with strong median vertical keel.
Suspensorium ( Fig. 5k, l View Fig ). Hyomandibula broad and deep, sutured to preopercle via lateroposterior flange, and metapterygoid via wide anterior process; anteriorly sutured and posteriorly synconchondrally jointed to quadrate. Cranial articulation of hyomandibula including slender anterodorsal process abutting weak ridge on sphenotic in front of hyomandibular facet; long, gently rounded condyle articulating with hyomandibular facet of sphenotic and pterotic; plus short, vertically-truncate posterodorsal surface articulating with pterotic behind hyomandibular facet. Lateral face with low, oblique rise between anterior process and preopercular flange, marking attachment limit of inner bundles of adductor mandibulae muscle. Low crest on medioposterior edge ventral to pterotic articulation, otherwise no enlarged articulating processes or muscle origin crests dorsal to opercle condyle. Opercle condyle centered slightly above midpoint on posterior margin; condyle twice as deep as wide. Lateral foramen of facial canal centrally located on anterior surface of adductor muscle crest at level of opercle condyle; medial foramen of facial canal anteriorly located above adductor arcus palatini crest. Medial face with centrally located vertical and crescentic adductor arcus palatini scar more prominent than in modern P. hemioliopterus ( Fig. 5m View Fig ) but is shape and placement similar.
Preopercle sutured to quadrate in addition to hyomandibula; lateral face shallowly concave forming fossa for posterior sections of adductor mandibulae muscle; posterior margin raised in a gentle curve and likely with sensory canal but no discernable lateralis pores; no evidence of external foramen for symplectic canal, but medial foramen of symplectic canal present between quadrate and preopercle.
Quadrate lateral face mostly shallowly concave; anteroventral blade broadly sutured to metapterygoid; mandibular condyle broad and strongly bilobed flanking central saddle, medial lobe of condyle braced by vertical buttress.
Anterior vertebrae ( Fig. 3b View Fig ). First centrum articulated to basioccipital and deeply sutured to compound or Weberian complex centrum (2-4). Aortic groove open along midventral line, flanked by low parallel ridges along first and compound centra; broken before centrum of vertebra 5. Weberian complex lacking prominent mid-dorsal vertical lamina; neural arch-spine complex incompletely preserved but anteriorly projecting to contact supraoccipital and exoccipitals. Indistinct pieces of tripus and low os suspensorium remain in place; anterior limbs of transverse processes meet compound centrum at right angle, wide and thickened laterally, broadly contacting ventral articulation flange of supracleithrum; vertebra 5 indeterminate.
Pectoral girdle ( Figs. 5 e, f, g View Fig ). Dorsal articulating process of cleithrum bifid, anterior limb longest, and overall similar in size to postcleithral process; postcleithral process deep and nearly equilaterally triangular, coarsely ornamented especially along ventral and ventrolateral edges lateral to articulating fossa of pectoral spine. In ventral view outward bulge of cleithrum in transverse alignment with posterior limit of articulating fossa of pectoral spine. Mesocoracoid not preserved but elevated surface near dorsal edge of coracoid shows its articulation site. Coracoid keel strongly elevated proximally, extending about midway to pectoral symphysis; coracoid keel divides jointed horizontal limbs of cleithrum and coracoid into equal halves; two parallel ridges run toward midline horizontal limbs of coracoid.
Pectoral spine shaft ( Figs. 5 e, f, h View Fig ) depressed, in section more ovoid than quadrangular, mostly covered by coarse, elongate and anastomosing ridges and grooves (neither meshlike nor finely-striate); anterior dentations erect, blunt, none strongly antrorse, relatively heavy and about half as numerous as posterior dentations; posterior dentations erect and irregularly spaced, some transversely widened but not exceptionally flattened, sharper and fewer than anterior dentations; anterior and posterior dentation rows without deep trenches and flanking ridges; no anterior distal serrations; spine tip bluntly pointed; spine base robust but details indeterminate.
| USB |
Bacterial collection of Universita degli studi della basilicata, Dipartimento di Biologia |
| FU |
Fudan University, Department of Biology |
| R |
Departamento de Geologia, Universidad de Chile |
| UNEFM |
Universidad Experimental Francisco de Miranda |
| MCZ |
Museum of Comparative Zoology |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phractocephalus nassi
| Lundberg, John G. & Aguilera, Orangel 2003 |
Phractocephalus hemiliopterus
| Lundberg, J 1997: 80 |
| Lundberg, J & Linares, M 1988: 131 |