
Bythotrephes lilljeborgi, Korovchinsky, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4379.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:EEF541E1-F91C-411E-9B65-2C79ED2A2706 |
|
DOI |
https://doi.org/10.5281/zenodo.5962980 |
|
persistent identifier |
https://treatment.plazi.org/id/60573E65-D160-FFAF-6EC2-67CFD0EE0674 |
|
treatment provided by |
Plazi |
|
scientific name |
Bythotrephes lilljeborgi |
| status |
sp. nov. |
Bythotrephes lilljeborgi sp. nov.
( Figs. 15 View FIGURE15 , 16 View FIGURE 16 )
Byhtotrephes longimanus s. str.: Lilljeborg 1901: 604 ‒617, Tab. 80, fig. 10‒19, Tab. 81, fig. 1‒2, Tab. 82, fig. 1‒10.
Etymology: The species is named in honor of the outstanding cladocerologist Prof. W. Lilljeborg (Uppsala, Sweden) who first described this species under the name “ Bythotrephes longimanus Leydig ”.
Type locality: Lake Ringsjön (Prov. Skåne, Sweden).
Type material. Holotype. A female with body length 2.08 mm from sample (Lilljeborg catalogue N 2678) labeled “ B. longimanus s. str., Sw, Sk, Ringsjön, 26.7.1876, coll. Lillj.” preserved and deposited in Museum of Evolution of Uppsala University (Uppsala, Sweden) (UPSZTY 167710).
Paratypes. All specimens, except holotype, enumerated below. They are deposited in Museum of Evolution of Uppsala University (see numbers below), Museum für Naturkunde (Berlin, Germany), Zoological Museum of Moscow State University (Moscow, Russia) and author’s collection.
Material examined. Sweden: 1) sample (MEUU, Lilljeborg catalogue N 2326) labeled “ Bythotrephes longimanus Leydig, Sw, Sk, Ringsjön , 26.7.1876, Lillj.”, 6 ad (paratypes, UPSZTY 167711); 2) sample (MEUU, Lilljeborg catalogue N 2606) labeled “ B. longimanus, Sw, Sk, Ringsjön , 24.7.1876, coll. Lilljeb.”, 15 ad (paratypes, UPSZTY 167712); 3) a slide (MEUU, Lilljeborg catalogue N P2329) labeled “ Bythotrephes longimanus ? Lillj., Sw, Sk, Ringsjön, 27.7.1878, Lillj.”, 2 ad, dissected (paratypes, UPSZTY 167713); 4) a slide (MEUU, Lilljeborg catalogue N P2330) with the same label, 2 ad, dissected (paratypes, UPSZTY 167714); 5) sample (MEUU, Lilljeborg catalogue N 2677) labeled “ B. longimanus s.str., Sw, Sk, Ringsjön, 26.7.1876, coll. Lillj.”, 3 ad, 1 juv. (paratypes, UPSZTY 167715); 6) sample (MEUU, Lilljeborg catalogue N 2678) with the same label, many ad (paratypes, UPSZTY 167716), a holotype was selected from this sample); 7) a slide (MEUU, Lilljeborg catalogue N P2325) labeled “ Bythotrephes longimanus Leydig, Sw, Sk., Ringsjön , 15.9.1882, Lillj.”, 2 males, dissected (paratypes, UPSZTY 167717); 8) a slide (MEUU, Lilljeborg catalogue N P2328) with the same label, 3 males (paratypes, UPSZTY 167718); 8) sample (MEUU, Lilljeborg catalogue N 2676) labeled “ B. longimanus, Sw., Sk., Ringsjön , 15.9.1883, coll. C. Möller”, 6 gam, 5 males, 1 juv. male (paratypes, UPSZTY 167719); 9) bottle (MNB, N 18911 View Materials ) with two tubes, the larger one labeled “ Bythotrephes longimanus , Skane, Ringsjön, 22 VI 97, Lilljeborg”, many ad (paratypes, ZMB 18911).
Norway (leg. T. Jensen): 1) Lake Holmsjøen , 7.7.2014, 3 juv. ; 2) Lake Storfiskevann , 6.8.2014, 4 ad, 2 juv. ; 3) Lake Sølensjøen , 18.9.2014, 2 ad, 3 males ; 4) Lake Sannes-Langen , 24.9.2014, 2 males .
Russia: 1) Nezhnekamskoye reservoir (55°42.511 N; 52°17.699 E), 20.8.2016, 4 ad, coll. V.I. Lazareva. GoogleMaps
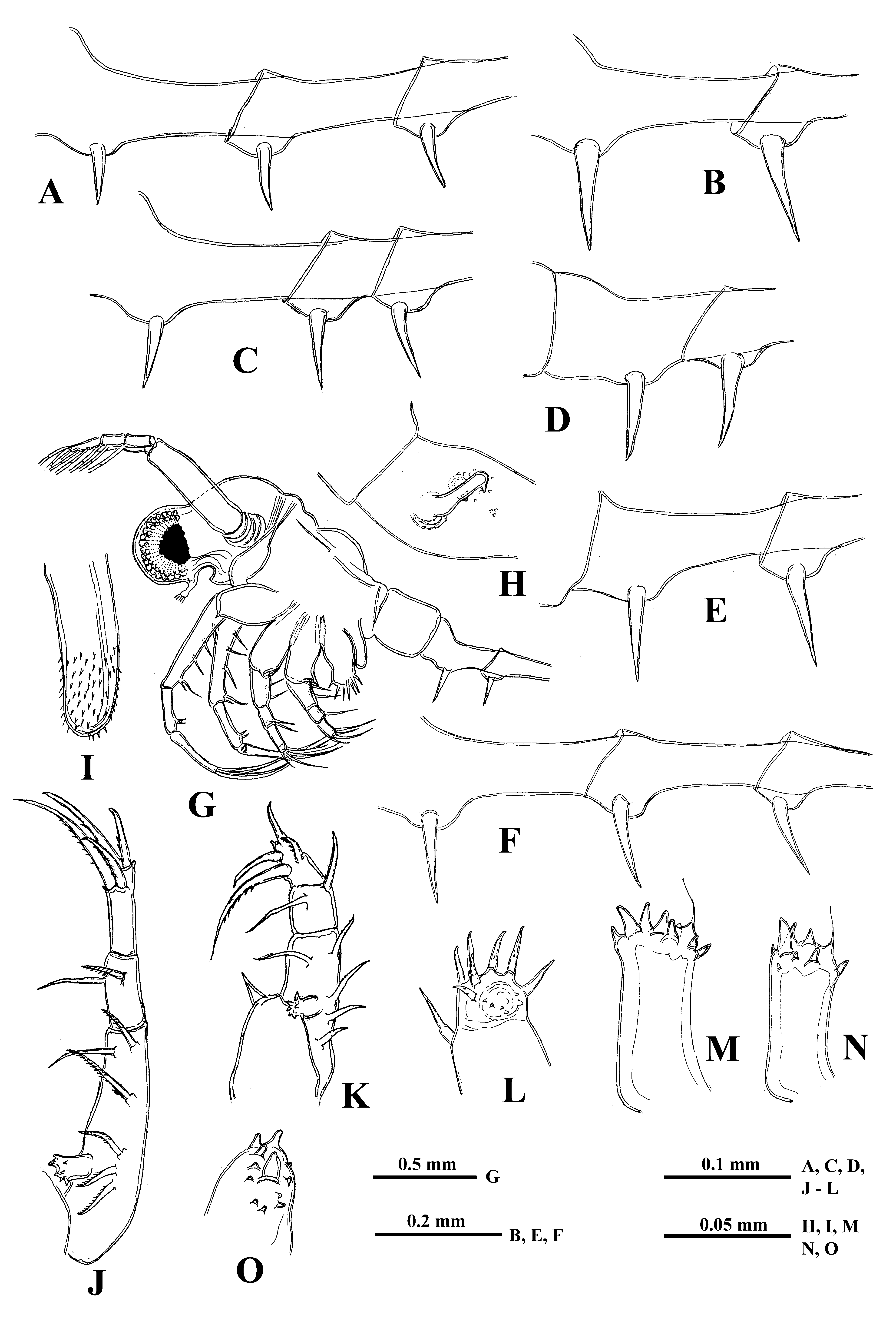
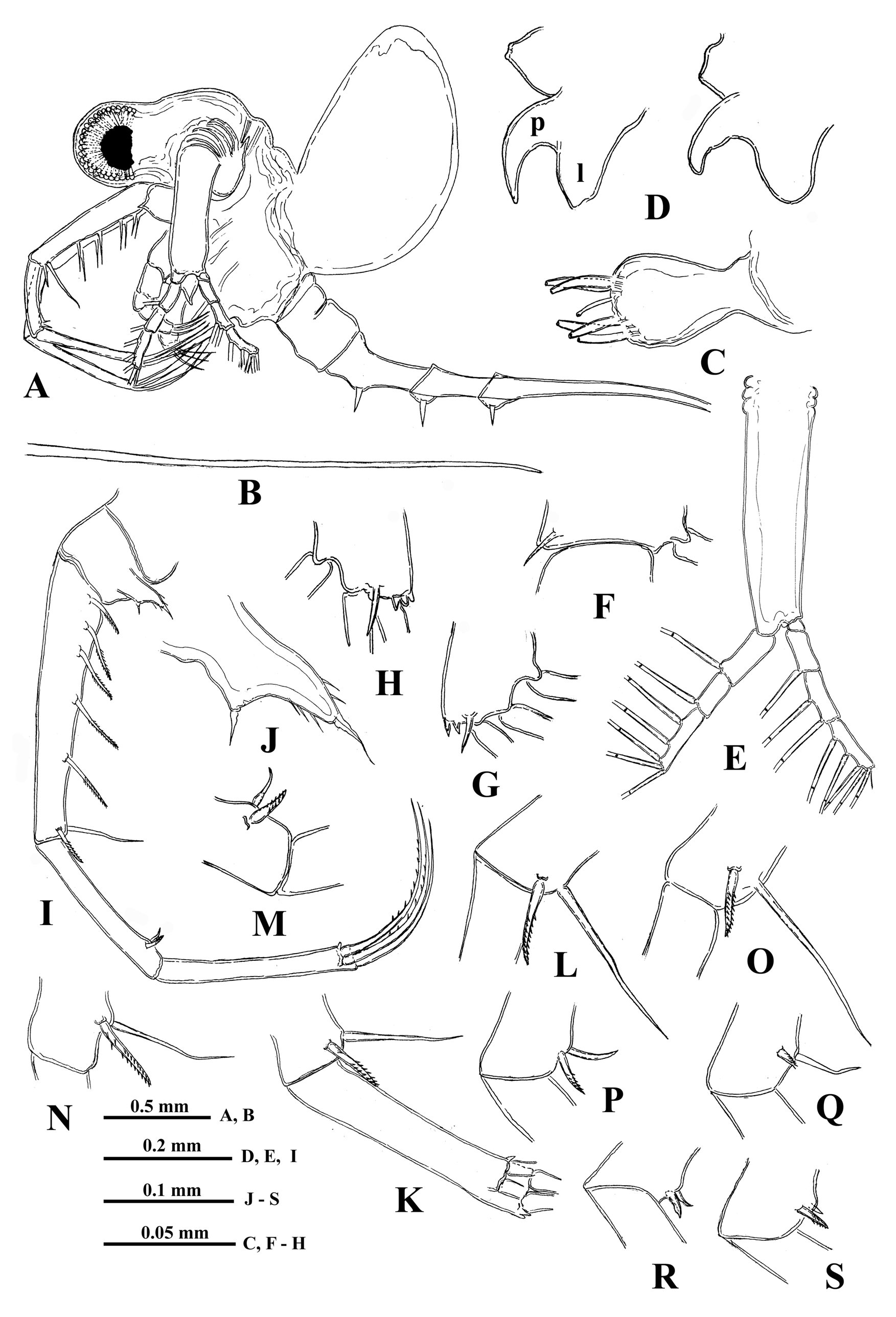
Description. Female. General body appearance and segmentation. Body elongated and divided into four parts: head, thorax, abdomen, and postabdomen with long caudal process ( Figs. 15A, 15B View FIGURE15 ). Its longitudinal axis is conspicuously incurved when the head is located at almost right angle to the thorax. Also a highly movable abdomen can be either in a straight line with the thorax or at different angles to it. Head large with rounded anterior part filled by the enormously developed compound eye and bearing small antennules ventrally. Posterior part of head bears long swimming antennae and mouth parts consisting of mandibles, maxillules (mx I), and upper lip (labrum). Thorax with strongly developed muscular ventral side bearing four pairs of thoracic limbs of different size directed antero-ventrally. Dorsally, thorax bears sack-like carapace transformed into a brood pouch, sometimes reaching large size. Abdomen (metasome) elongated, cylindrical, inconspicuously three-segmented (see Korovchinsky 2015) and flexible, connected with small postabdomen, bearing ventrally a pair of straight claws and posteriorly very long straight caudal process with either one or two pairs of similar claws proximally. General body length of females (without caudal process) may reach 2.3 mm or slightly more (in the examined specimens it ranges from 1.41 to 2.37 mm) while the length of caudal process may exceed the body length by 1.4‒2.4 times (mean = 1.9 times).
Head. Comparatively large (38.1‒42.2 % of body length) and subdivided into two parts: rounded anterior part mostly filled by large compound eye and posterior part bearing dorsally a large saddle-shaped organ, swimming antennae and mouth parts. Large pigment spot occupies about one-third or at most a half of the eye’s volume. Ocellus (naupliar eye) is absent (see Elofsson 1966).
Antennules. Small and situated on the ventral side of the anterior head part beneath the eye. They are bulbous ( Fig. 15C View FIGURE15 ) and sit on the joined basis slightly split anteriorly. Terminally they bear five regular aesthetascs in two groups of three and two, and one shorter and thinner aesthetasc-like structure, situated in a group with two regular aesthetascs, and having slightly widened apical end (“accessory simple seta” according to Scourfield (1896)).
Swimming antennae. Comparatively long, with elongated cylindrical basipodite ( Figs.15A, 15E View FIGURE15 ). Of two antennal branches, the lower three-segmented one (endopodite), sitting on the apical basipodital prominence, is slightly longer than upper branch. The upper branch is four-segmented and lower branch is three-segmented ( Fig. 15E View FIGURE15 ). Proximalmost segment of the upper branch is rudimentary and clearly visible only externally, all other segments of both branches are much more developed. Small spine on the end of second segment of upper antennal branch ( Fig. 15F View FIGURE15 ); similar apical spines and tiny denticles on the distal segments of both branches ( Figs. 15G, 15H View FIGURE15 ). Small proximalmost segment of upper branch lacks setae, while other segments possess two-segmented swimming setae of more or less similar size except distalmost of them which are shorter. All setae bilaterally armed with rows of uniform thin setulae. General formula of antennal setae: 0‒1‒2‒5/1‒1‒5.
Mouth parts. They are represented by upper lip (labrum), mandibles, and maxillules (maxilla I). The upper lip is composed of two parts: the posterior thick and slightly flattened triangular lobe and anterior large proboscis-like outgrowth ( Fig. 15D View FIGURE15 ). The latter is separated from the former one by a deep indention of the cuticle. Mandibles and maxillules (mx I) are similar to those of previously described species.
Carapace. It looks like a bag-like structure, strongly modified into closed brood pouch ( Fig. 15A View FIGURE15 ) widely connected in its base with dorsal side of thorax. It may be often well developed being filled either by eggs or large embryos.
Thoracic limbs. Four pairs of strongly chitinized, stenopodous limbs are densely situated along the muscular ventral side of thorax and directed antero-ventrally ( Figs. 15A View FIGURE15 , 16G View FIGURE 16 ). All of them have complex and variously setaceous armament along their inner side. Limbs of three anterior pairs are five-segmented and those of the last fourth pair are three-segmented. Protopodites of all of them, covered by comparatively softer cuticle, are inconspicuously delimited into two parts (segments), coxa and basis, while the endopodites of limbs of three anterior pairs are composed of three well developed segments and those ones of the fourth pair are unisegmented ( Figs. 15I View FIGURE15 , 16J, 16K View FIGURE 16 ).
Limbs of first pair (tl I) are especially long and strong, their length can be rarely almost equal to the length of the body (up to 97.6 % of its length) but usually they are shorter (58.7‒88.4 % of body length) ( Fig. 15I View FIGURE15 ). Terminally, the inner side of their protopodite bears a small triangular lobe, the pseudognathobasic process (see the explanation of the term in Korovchinsky (2015)), armed laterally and distally with two outgrowths with apical setae and numerous spinules ( Fig. 15J View FIGURE15 ). The external part of protopodite is longer than the internal one and bears apically a small conical outgrowth. The first segment of the endopodite is long and bears 5‒6 anterior lateral setae (their number on limbs of one individual sometimes can vary) with a posterior row of rough incurved spines and anterior and lateral rows of fine setulae. Distally, this segment bears a shorter anterior seta of the same type and a long posterior finely setulated seta ( Figs. 15L, 15O View FIGURE15 ). The second segment of the endopodite is conspicuously shorter and bears only two apical setae similar to those on the end of previous segment but usually shorter. Its size can vary considerably ( Figs. 15M, 15N, 15P, 15Q, 15R, 15S View FIGURE15 ), however, the rudimentary setae ( Figs.15R, 15S View FIGURE15 ) are present very rarely. The terminal third segment of endopodite varies in length but generally it is comparatively short (20.0‒29.3 % of body length) and always bears apically four long roughly spinulated setae, two of them terminally and two subterminally ( Figs. 15I, 15K View FIGURE15 ). Basally, these setae are armed by a row of smaller spines, while distally, by larger lanceolate spines situated in two rows and directed terminally.
The limbs of the second pair (tl II) are considerably shorter, their protopodite, again externally, is conspicuously longer and bears a conical outgrowth. The first, basal segment of their endopodite, bears a row of 4‒ 6 rather long anterior lateral setae ( Fig. 16J View FIGURE 16 ). Also there may be one posterior lateral seta of the same type on this segment. The terminal setae of the segment are different, the anterior one is shorter and roughly armed, while posterior one is longer and finely setulated. Internally, this segment bears stout cylindrical pseudognathobasic process, possessing some prominences of different size and one small, thin seta ( Fig. 16M View FIGURE 16 ). The second segment of endopodite is short with only two setae, the anterior of which is similar to anterior terminal seta of previous segment, while the posterior seta is longer and finely setulated. The distal, third segment of endopodite of the limb bears four setae, two terminal and two subterminal. Their armament is similar to those of the previously described species.
The limbs of the third pair (tl III) ( Fig. 16K View FIGURE 16 ) are generally similar to those of the previous ones, differing in some details. The external outgrowth of their protopodite is conspicuously larger and lateral anterior setae of first segment of endopodite are fewer (3‒5) while the posterior ones are absent. Distal setae of the segment are similar to other ones. The pseudognathobasic process is also similar to that one of tl II ( Fig. 16N View FIGURE 16 ). Of setae of the second segment, the anterior one is similar to the respective one of tl II. Terminal and subterminal setae of third segment are similar to those of tl II but slightly shorter bearing fewer denticles.
The limbs of the fourth pair (tl IV) are considerably reduced; their protopodite bears slightly spinulated seta sited on a short cylindrical base ( Fig. 16L View FIGURE 16 ). The only segment of endopodite has two rows of comparatively short spine-like setae. The external row (group) always consists of two setae, and the internal row of 5‒7 setae, differing in their appearance and armament. Almost the whole internal part of the endopodital segment is occupied by the reduced pseudognathobasic process, also armed by some denticles and thin seta ( Fig. 16O View FIGURE 16 ).
Abdomen (metasome) is often deformed. It is inconspicuously delimited in two segments, short proximal and long distal with prominent fold more or less in the muddle dorsal side.
“ Postabdomen”, actually consisting of two parts, the last small abdominal segment and postabdomen per se (see Korovchinsky (2015)), is comparatively small; the anal opening is situated between postabdominal claws. The latter ones are of moderate size (5.4‒12.1 % of body length), usually straight and directed downwards, sometimes with apex curved slightly forward ( Figs. 16A, 16B, 16C, 16D, 16E, 16F View FIGURE 16 ).
Caudal process is directly connected with postabdomen and proceeds as a very long, proximally rather thick and curved, then straight spine-like structure ( Figs. 15A, 15B View FIGURE15 ) variable in its length (142.3‒241.0 % of body length), thus surpassing the body length about in one and a half - two and a half times. Basally, caudal process bears one or two pairs of claws similar to those of postabdomen (e.g., proximal claws reach 6.1‒11.4 % of body length) ( Figs.16A, 16B, 16C, 16D, 16E, 16F View FIGURE 16 ). Pairs of claws usually sit rather closely (e.g., distance between postabdominal claws and proximal claws of caudal process (interclaw distance) constitutes 9.2‒19.3 % of body length). Between the latter, the thickness of the structure constitutes 4.0‒7.3 % of body length. Borders separating old molted integuments of caudal process with claws normally are quite conspicuous.
Size 1.41‒2.37 mm, Lilljeborg (1901) recorded the body length up to 3.0 mm.
Gamogenetic females differ from parthenogenetic ones only in presence of large yellow-brownish resting eggs (just forming ones reaching 0.32 mm in diameter) in their brood pouches.
Juvenile females. The investigation of few juvenile females (with only a pair of postabdominal claw) (body length 1.47‒1.57 mm) has revealed that they differ from adults in presence of somewhat longer thoracic limbs of first pair (tl I) (85.3‒92.4 % of body length) as well as longer (283.0‒319.0 % of body length) and basally thicker (7.1‒7.6 % of body length) caudal process.
Males. Males ( Fig. 16G View FIGURE 16 ) have body size more or less similar to that of females. Thoracic limbs of first pair (tl I) are comparatively short (61.3‒76.7 % of body length) as well as each segment of them, especially distal one (16.3‒19.1 % of body length), which is slightly swollen proximally and bears on its inner side a small strongly chitinized hook with two inner denticles; a field of tiny prominences is situated under it ( Fig. 16H View FIGURE 16 ). The copulatory appendages are small and armed with numerous minute spinules terminally ( Fig. 16I View FIGURE 16 ). Body length 1.95‒2.56 mm.
Remarks. Lilljeborg (1901) presented rather detailed description of the species under the name “ B. longimanus ”, though it contained a number of errors, in particular in the description of structure of the thoracic limbs, abdomen, and postabdomen. Generally, the specimens of the taxon investigated by the author, if compared with other forms of Bythotrephes , mostly reminded B. longimanus s. str. which by that time was not described and depicted well enough (see Korovchinsky (2015)). Due to purely regional character of Lilljeborg’s studies which lacked detailed interregional comparisons, the taxonomic identification of the material under consideration turned out to be incorrect.
In the course of his survey of “ B. longimanus ”, Lilljeborg (1901) also recorded a form from Lake Mycklaflon (Småland, Sweden) having an unusually long and distally thin tl I with short apical setae on two proximal endopodital segments reminding those of B. longimanus s. str. Such a material was not found in the course of present investigation, though the specimens of a new species from Lake Ringsj ӧn also had rather long tl I with comparatively short apical setae of second segment of their endopodite. Thus, potentially the Mycklaflon specimens can belong to B. lilljeborgi sp. nov. On the other hand, it may be suggested that they may represent the long-limbed representatives of B. brevimanus (see above).
After Lilljeborg’s (1901) description, there was no hint of finding this species anywhere else. Similar features may be suggested in poorly described individuals from England (see Beck 1883) which were incorrectly assigned to B. longimanus s. str. (see Korovchinsky 2015). In another case, Agnesotti (1935) recorded similar specimens with one-two pairs of claws on caudal process from Lago Lario (Lake Como, Comersee) in Italy (“ B. longimanus lariano ”) although other researches ( Ischreyt 1939, Korovchinsky 2015) found here B. longimanus s. str. with just one pair of claws of caudal process. Agnesotti (1935) found that the features of his specimens had much in common with those of Lilljeborg’s (1901) “ B. longimanus ”. At the end, a form of Bythotrephes with similar claws was found in Nizhnekamskoye reservoir ( Russia). The taxonomic status of all these forms is unclear and requires further resolution.
Differential diagnosis. A new species under consideration differs from B. longimanus s. str. first of all in presence of two-three pairs of claws in adult females instead of presence of not more than two pairs in the latter species. Also it possesses shorter tl I with usually shorter distal segment and larger setae on distal ends of first and second segments of their endopodites. From B. brevimanus , it differs in presence of larger claws of postabdomen (5.4‒12.1 (av. 6.6‒9.3) % vs. 0.8‒7.8 (av. 2.4‒6.3) % of body length) and caudal process (6.1‒11.4 (av. 6.9‒9.3) % vs. 2.3‒8.3 (av. 3.1‒6.4) % of body length) which normally directed more downwards.
B. arcticus possesses considerably larger body size, basally thicker caudal process, and its claws are stouter and usually curved backwards. B. cederströmii possesses even more prominent differences having very long caudal process usually supplied with denticulated bend and its postabdominal and caudal claws are much larger, curved forward apically and inserted distantly from each other.
For clearer species recognition, the identification key for the known species of the genus is provided (for better identification, a number of specimens from any population, not single ones should be used).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SuperOrder |
Cladocera |
|
Order |
|
|
Family |
|
|
Genus |
Bythotrephes lilljeborgi
| Korovchinsky, Nikolai M. 2018 |
Byhtotrephes longimanus
| Lilljeborg 1901 : 604 |