
Myotis annatessae, K Ruskop & B Orisenko, 2013
|
publication ID |
https://doi.org/ 10.3161/150811013X678937 |
|
DOI |
https://doi.org/10.5281/zenodo.4323722 |
|
persistent identifier |
https://treatment.plazi.org/id/5E5B87D0-1D4A-732C-FC60-7C65E078FE58 |
|
treatment provided by |
Carolina |
|
scientific name |
Myotis annatessae |
| status |
sp. nov. |
Myotis annatessae View in CoL sp. nov.
Holotype
ZMMU S-165042 (alcohol-preserved, skull extracted), adult male; Song Con river valley , Saola Basecamp, Vu Quang, Ha Tinh Province, Vietnam. 30.07.1997; coll. A. V. Borisenko.
Paratypes
Four adult females, ZMMU S-164986, 164988, 165046, 165047, four adult males S-164987, 164989, 165043, 165044 (alcohol-preserved, skulls extracted), 30.07– 27.08.1997, same locality and collector.
Referred material
Two adult females, ZMMU S-165051-165052 (alcohol-preserved), same locality as type; one adult female, ROM MAM 106476 , Nam Pan, N. of Lac 20, Khammouan Province, Laos, 20.04.1996; coll. C. M. Francis.
Type locality
Song Con river valley, Vu Quang, Ha Tinh Province, Vietnam.
Diagnosis
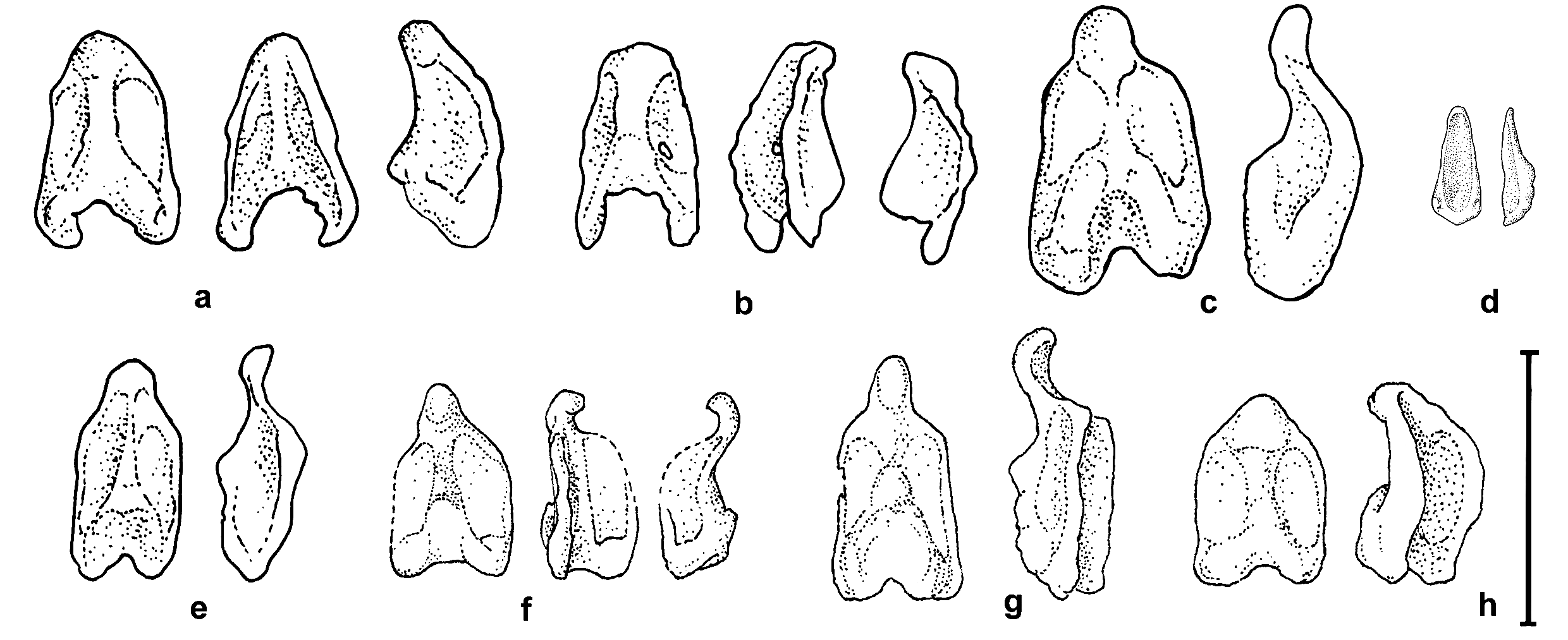
Small bat, similar in external, cranial and dental qualitative characters to M. muricola and to Myotis mystacinus sensu lato. Differs from M. muricola and M. nipalensis by smaller overall size, cranial dimensions and greatly reduced upper and lower canines, similar to those of M. siligorensis . Differs from M. siligorensis by a less concave frontal profile of the skull and myotodont lower molars. Differs from M. muricola by longer thumb and proportionally larger foot. Baculum is saddle-shaped but in details distinct from that of all the Indochinese mystacinus -like Myotis and bats of M. siligorensis lineage ( Fig. 3 View FIG ).
DNA barcode of the holotype
GenBank accession JF443982 View Materials , BOLD process ID ABBM488-07. DNA barcode sequence divergence from nearest neighbour is ca. 6%.
Measurements of the holotype (in mm)
External measurements: head & body 38, tail 37, forearm 34.3, hind foot 6.2, tibia 14.5, thumb (without claw) 5.3, ear pinna 13.2, wing span 230. Skull measurements: CBL 12.37, CCL 11.53, OH 5,22, MW 6.98, BCW 6.78, IOW 3.42, RW 3.52, RL 2.70, CM3 4.80, P4M3 3.52, C 0.62, Pseud 0.61, CC 3.41, M3M3 5.24, M2 1.25, MdLg 9.35, MdLa 9.57, cm3 5.09, p 4m 3 3.72, MdH 2.38.
Etymology
The species epithet is coined in honour of AVB’s daughter, Anna Tess.
Description
Small-sized Myotis (FA ca. 32.6–35.3 mm, BM ca. 2.9–4.3 g.) similar in appearance to M. muricola or M. mystacinus . Tail shorter than head and body by about 5 mm. Fur not particularly thick, hairs relatively short (ca. 4–6 mm) extending by 4–5 mm onto the tail membrane. Pelage coloration greyishbrown on back and paler on belly. Individual hairs on back are blackish at bases gradually turning brown towards the tips; ventral hairs more contrastingly tricoloured, with blackish bases, brown midportion and silvery-whitish tips. Genital area sparsely covered with white hairs. Hairless parts of skin mid-brown. Muzzle also covered by sparse
hairs almost up to the nostrils. Ears narrow, moderately long, reaching to the tip of muzzle if laid forward, bluntly pointed at tips, with very weak posterior emargination. Tragus bluntly pointed, relatively short, not reaching half of ear conch in height. Wing moderately long, with third metacarpal definitely longer than forth and fifth. Thumb proportionally long, with long and sharply pointed claw. Hind foot measured without claws about 42% of tibia length. Calcar lobe poorly developed. Wing membrane attaches to the base of the outer finger. Penis is relatively small, but not thin, slightly bulbously widen- ed in distal half.
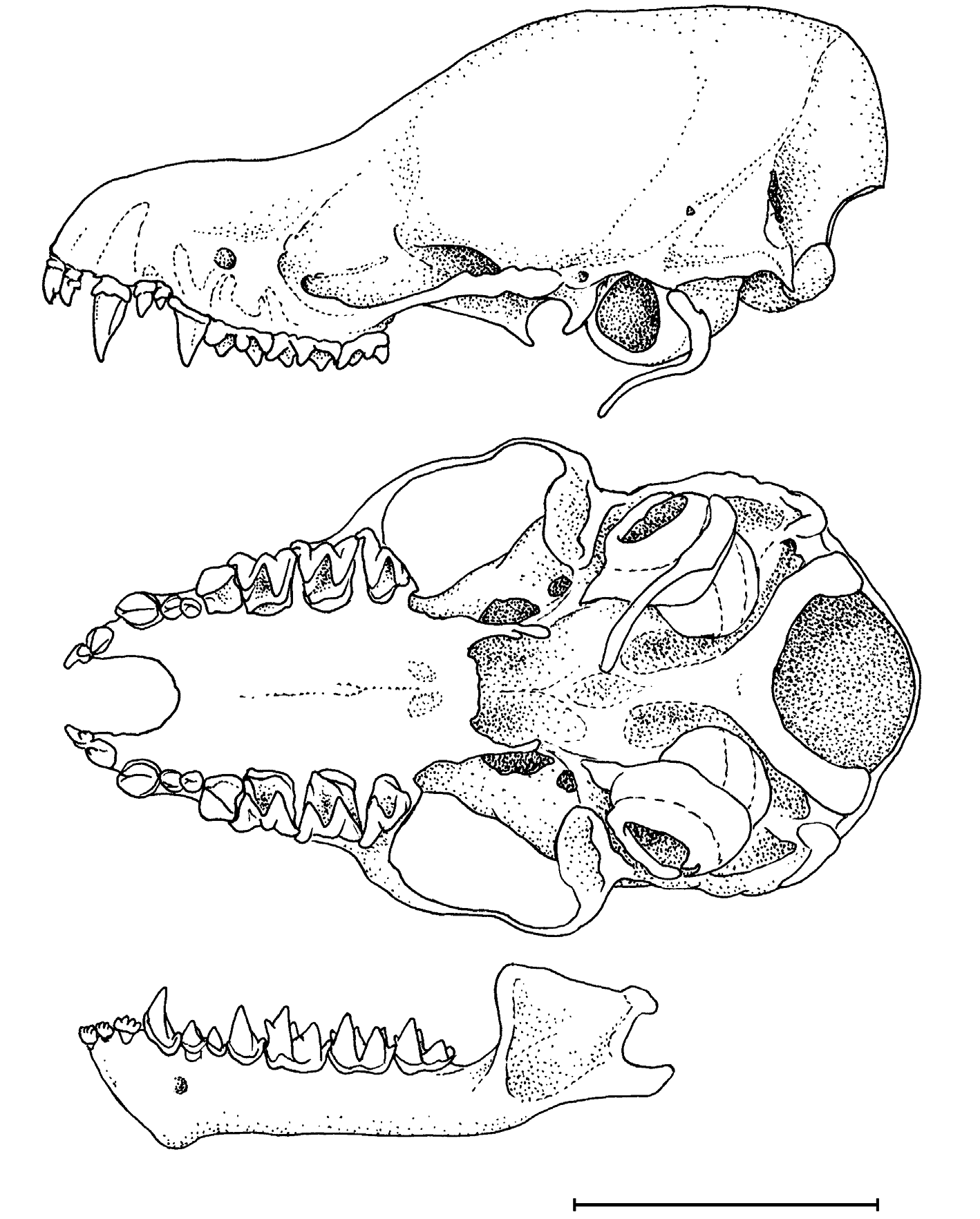
Skull small (average CCL 11.60 ± 0.24 mm; mean CM 3 4.79 ± 0.11 mm). In general appearance, the skull is intermediate between that of M. muricola and M. siligorensis : lightly built, with low and narrow rostrum (though more massive than in M. siligorensis ), somewhat bulbous braincase and definite, but not deep concavity on the frontal profile ( Fig. 5 View FIG ). In lateral view, highest part of skull is at the level of the supraoccipital bone. Sagittal and occipital crests absent, lambdoid crests poorly developed but visible. Zygomatic arch thin, without any vertical lobes. Anteorbital opening is situat- ed over the posterior root of P 4 or over the gap between it and the first root of M 1; maxillary channel moderately short. Posterior palatal emargination is rectangular in shape. Basicranial pits are well-developed. Lower jaw with almost vertical anterior edge of coronoid process; angular process definitely longer than articular, its distal end somewhat curved upward.
Teeth are relatively gracile. Canines (especially lower) are small, only slightly exceeding corresponding posterior premolars in height. Upper canines thin, almost round in occlusal view, with shallow but definite outer and inner posterior grooves. Outer upper incisors are about 2/3 of inner incisors in crown area and slightly smaller in height. Inner incisor clearly bicuspidate; outer incisor with a definite supplementary cusp on its lingual side. Inner lower incisors are with rudimentary fourth cusps; middle and outer lower incisors definitely four-cusped. Small upper and lower premolars positioned within tooth rows. Cingulum of the large upper premolar with a well pronounced anterior cusp. Upper molars with only traces of hypocones and paraconules, with closed trigon basins. Lower molars of myotodont type, with well developed hypoconulid; talonids on M 1–2 slightly larger than corresponding trigonids.
point of the lateral projection just in front of it, approximately in the basal third ( Fig. 3a, 3b View FIG ). Urethral groove is deep and wide. Distal end of baculum blunt, without any abrupt constrictions.
Comparison to other species
The new species mostly resembles M. muricola s. lato in general appearance. It differs in smaller average size (both in external and cranial measurements) and weaker dentition with particularly small upper canines. The thumb and its claw is longer than in Indochinese and Malayan M. muricola ; thumb measured without claw about 15% of forearm length (13.7–16.9 vs. 9.2–12.3% in M. muricola ). The calcar lobe is less developed than in M. muricola . The notch on the posterior edge of the ear conch is noticeably better developed in M. muricola . Penis is somewhat smaller and less bulbous than in Indochinese M. muricola and M. ater , but definitely thicker than in M. mystacinus s.lato. Indochinese specimens of M. muricola also have, on average, a proportionally shorter hind foot (about 38% of tibia length).
From M. ater , the new species also clearly differs in smaller size and lighter coloration. From M. nipalensis , it differs in a smaller upper canine and proportionally higher braincase: the average OH/MW ratio is 0.75 in M. annatessae , and 0.69 in M. nipalensis . From M. siligorensis and its allies the new Myotis differs by distinctively myotodont lower molars, larger canines (which exceed the corresponding large premolars in height), rectangular (not U or W shaped) posterior edge of palate and a more robust rostrum.
The new species also differs from other morphologically similar Myotis in baculum shape. In the M. siligorensis complex, the baculum is minute and peg-like, often lacking the urethral groove. In Indochinese M. muricola and M. ater it is clearly different, with a constricted distal end. Amongst all the specimens analyzed, the baculum of M. annatessae bears closest resemblance to that of the Nepalese M. muricola , but the latter is more robust, more curved upward and has very shallow basal concavity ( Fig. 3 View FIG , h).
From M. laniger and small individuals of M. horsfieldii , the new species is easily distinguishable by a smaller hind foot (about half of tibia length in both species and definitely shorter in M. annatessae ), by the place of the wing membrane attachment (above the base of the outer toe in both M. laniger and M. horsfieldii ) and by shorter canines; from M. rosseti — by larger foot, larger overall size, absence of the adhesive pads on thumbs and both small middle upper premolars well developed. According to published data ( Allen, 1938; Smith and Xie, 2008), M. davidii — another East Asian mouseeared bat of similar size — has a larger hind foot and posterior upper small premolars displaced inward from the tooth rows.
Several other named forms of small-sized Myotis have been described from adjacent parts of mainland South-East Asia and were traditionally treated as subspecies of M. muricola ( Simmons, 2005) . Myotis lobipes from western Burma has long been regarded as a full synonym of M. muricola s. str. (Tate, 1941). Its type specimen resembles M. muricola specimens from Indochina in size and qualitative features.
Myotis moupinensis from China (not included into our analysis) is also usually treated as a partial synonym of M. muricola (Tate, 1941; Simmons, 2005). Its average smaller size ( Allen, 1938) gives it superficial resemblance to M. annatessae . However, six alcohol-preserved specimens in ZISP, labelled as M. moupinensis and collected from within the known distributional range of this form, possess nyctalodont lower molars and other morphological traits in common with the Taiwanese M. latirostris . Thus they are more likely to be affiliated with this distinct species (M. Ruedi, personal communication). The same is probably true for M. blanfordi and M. caliginosus . Specimens of these forms (including types of M. blanfordi ) processed in the Berlin Zoological Museum match M. latirostris in cranial shape and dentition. This entire species complex, while similar in size to M. annatessae , differs markedly in having a lower braincase, shallower frontal profile, proportionally larger canines, and nyctalodont lower molars.
Finally, available skull measurements of the form niasensis, described from Nias Island as “a small form of Myotis muricola ” ( Lyon, 1916: 442) , resemble those of the new species, but niasensis has short- er forearm and ear lengths (31.2–31.5 and 9.8–10.2 vs. 32.6–35.3 and 11.6–13.8, respectively). Tate, who apparently saw the type of niasensis, reports that it has a low brain case, in contrast to the proportionally high brain case in M. annatessae .
Genetic analysis
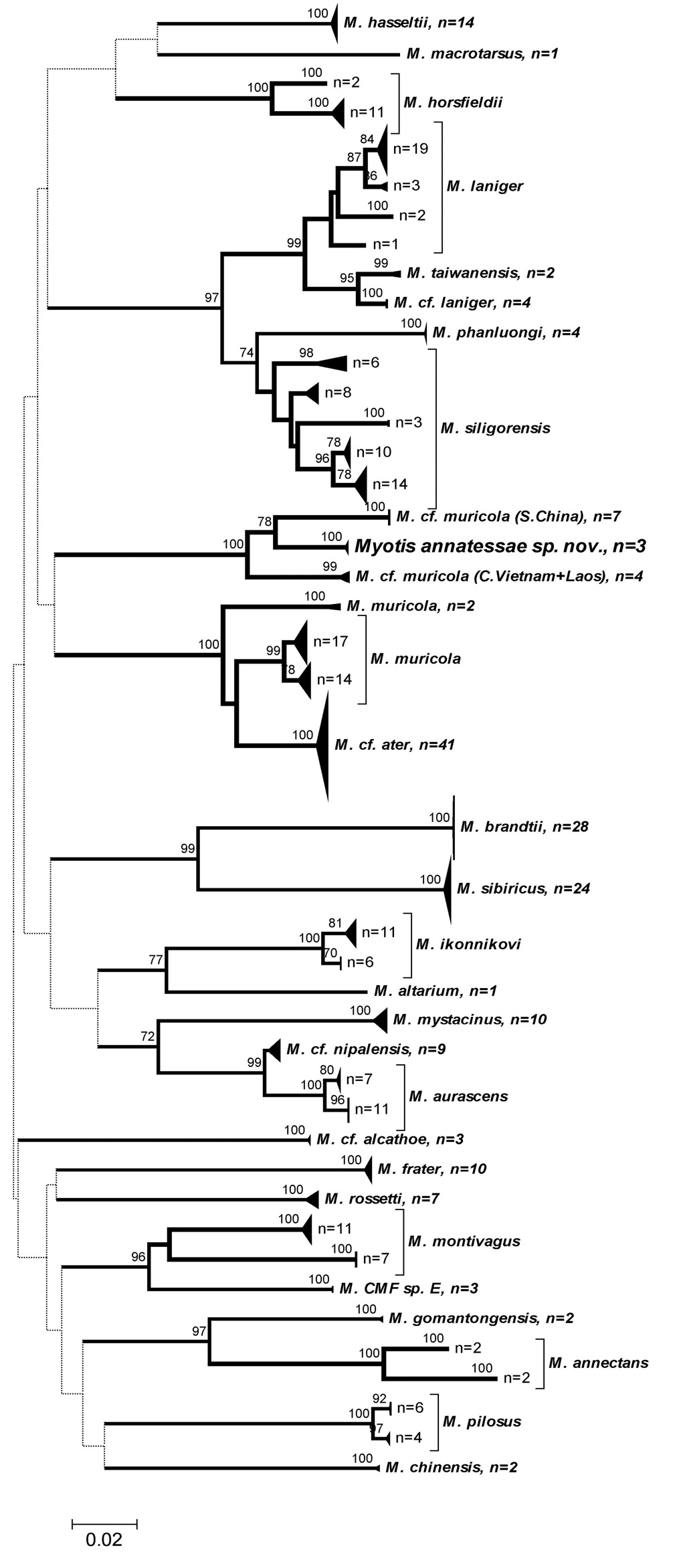
As mentioned earlier, neighbour-joining analysis of COI sequences ( Fig. 4 View FIG ) corroborates the observed morphological distinctiveness of M. annatessae from both M. muricola and M. siligorensis complexes, as well as from the extralimital M. mystacinus and allied species. Available genetic data are insufficient to infer phylogenetic relationships between the new species and other Myotis , but its deep genetic divergence from the most morphologically similar representatives of this genus is clear. Closely related lineages, labelled on the tree as ‘ Myotis cf. muricola ’, most probably represent undescribed forms. However their divergence from M. annatessae also corresponds to specific level.
Comments on natural history
All Vu Quang specimens in the type series were captured over the Song Con River. Bats were observed foraging about 1–7 m above the water surface, over ripples as well as over backwaters. In the latter case M. annatessae were observed togeth- er with M. horsfieldii , although the latter was foraging immediately above the water surface; thus two species partitioned the foraging space by altitude. Echolocation calls recorded with a QMC Mini heterodyning ultrasound detector were FM, low intensity, with frequency ranging from 45–50 kHz. All specimens captured in July–August were reproductively inactive.
Distribution
We captured this species only in the type locality. The species is also known from Khammouane Province in Central Laos (ROM MAM 106476). The Laotian record was made approximatly 40 km from the type locality. Morphologically and genetically similar specimens, though segregated by a relatively high genetic distance, were
collected in Ngoc Ling, Central Vietnam (ROM MAM 111270) and in Guizhou Province, China (ROM MAM 117808, 117809). Collecting localities were confined to large rivers in mountain foothills at elevations from 200 to ca. 1,300 m a.s.l.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |