
Arthropeina
|
publication ID |
https://doi.org/10.11646/zootaxa.3827.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:D6C800AA-8127-41D1-ACEA-52254F7CE89D |
|
DOI |
https://doi.org/10.5281/zenodo.6124288 |
|
persistent identifier |
https://treatment.plazi.org/id/5E0C3871-7543-FFCC-B0D0-6A53144FE08B |
|
treatment provided by |
Plazi |
|
scientific name |
Arthropeina |
| status |
|
Arthropeina View in CoL apomorphies
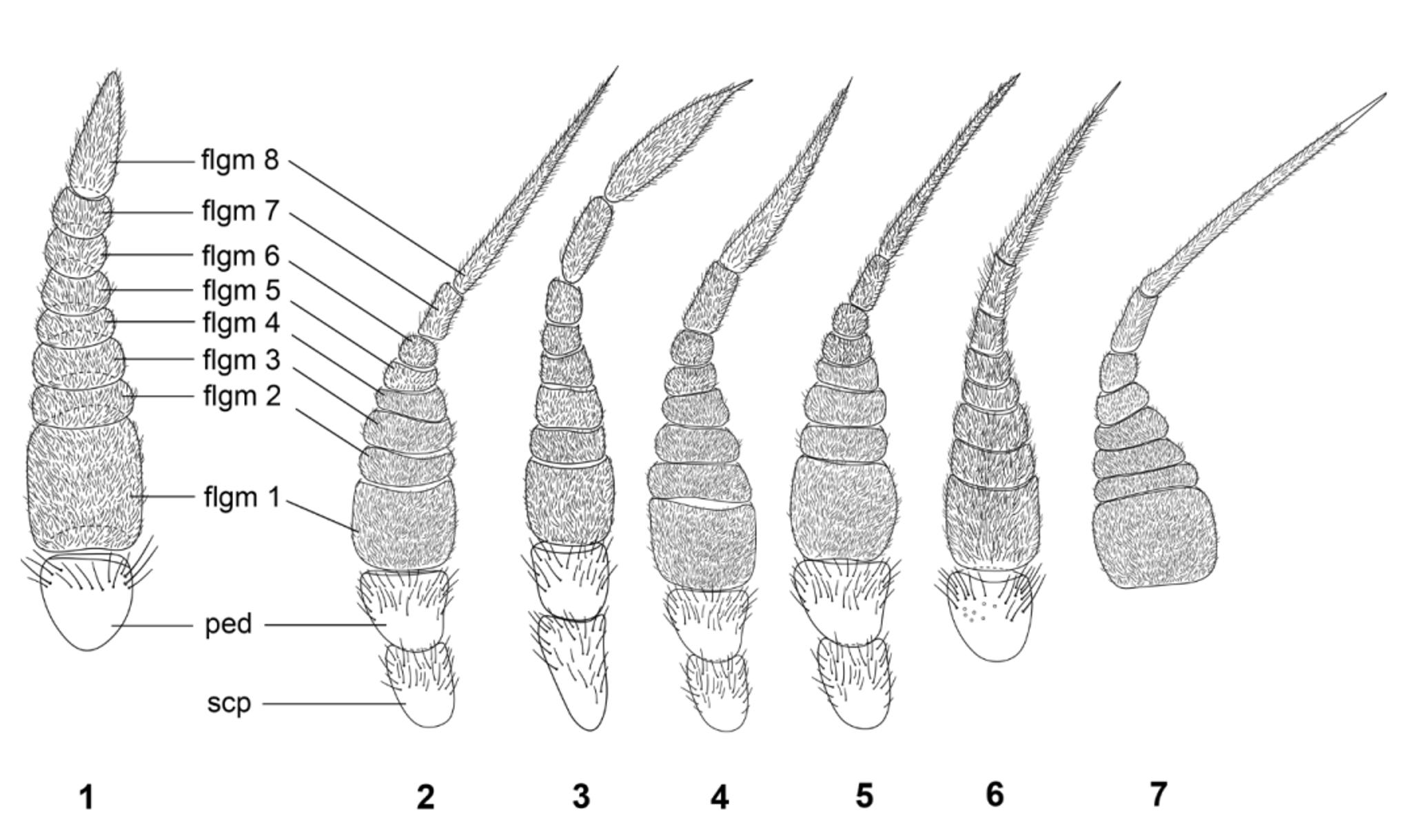
The conical shape of the antenna, with flagellomeres 7 and 8 together, forming a stylus ( Figs. 2–7 View FIGURES 1 – 7 ) that is as long as or longer than the remainder of the flagellum has been proposed as synapomorphic for the genus ( Artigas & Papavero, 1991; Woodley, 2011). This differs from the combined length of flagellomeres 7 and 8, which are shorter than the remaining flagellomeres in Solva and do not form a stylus ( Fig. 1 View FIGURES 1 – 7 ). In Arthropeina , the flagellomeres differ from each other in size and shape ( Figs. 2–7 View FIGURES 1 – 7 ), while in Solva ( Fig. 1 View FIGURES 1 – 7 ; Daniels, 1976, fig. 3) most flagellomeres are similar in size and shape (except for flagellomere 8 which is slightly narrowed towards apex). This is another feature that may be synapomorphic for Arthropeina . A more detailed comparison between additional species of Solva and other xylomyids would make this statement more robust.
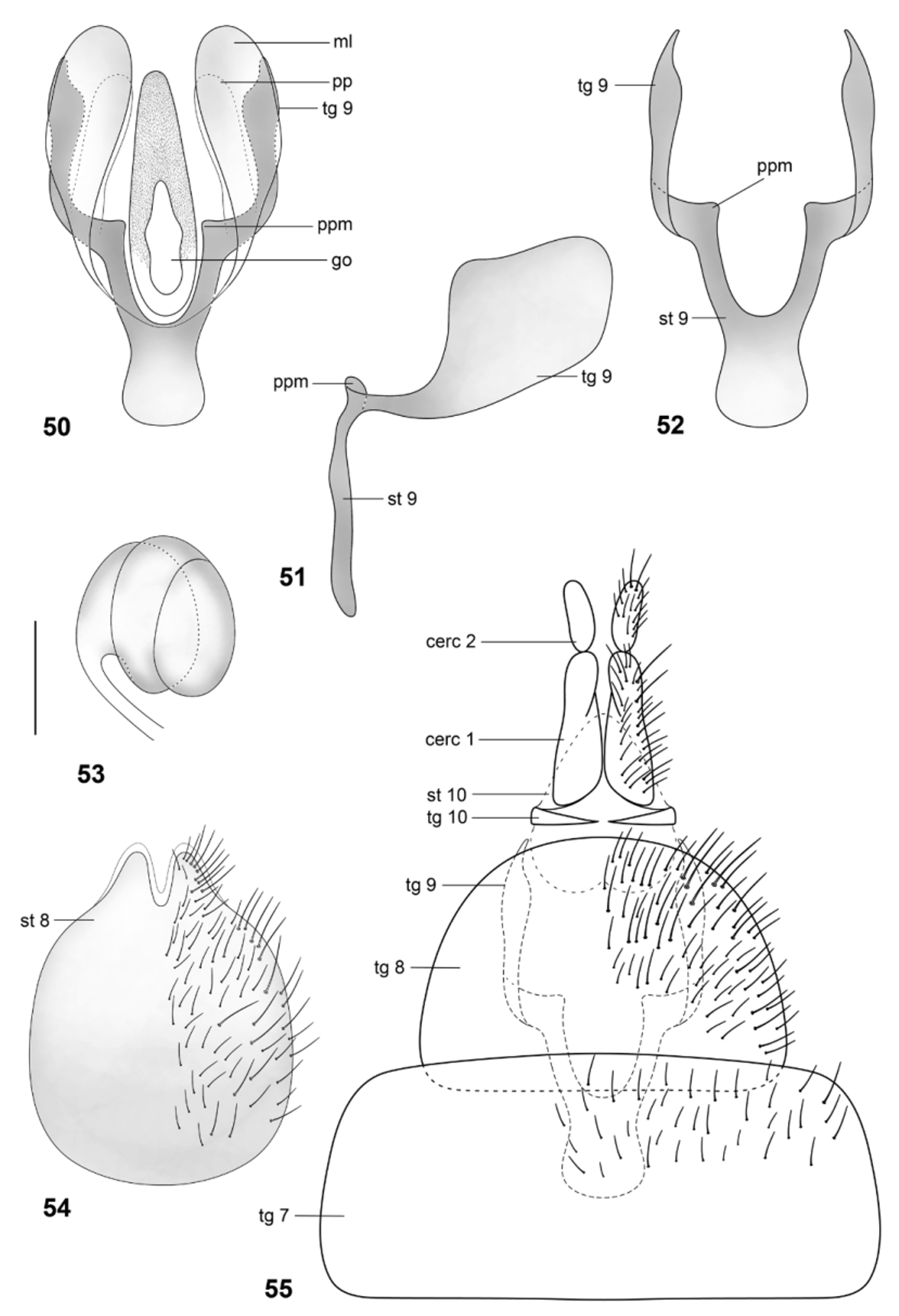
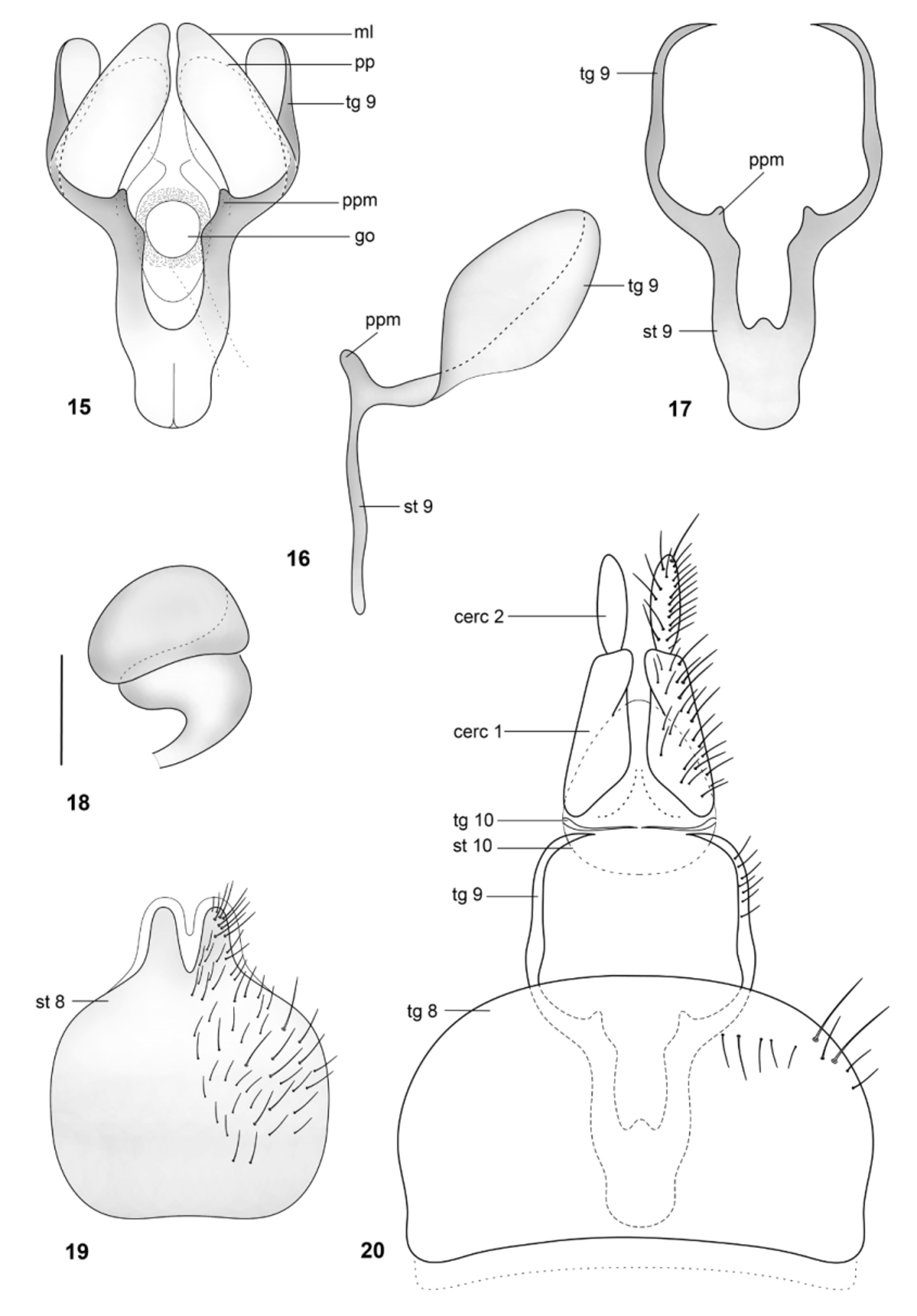
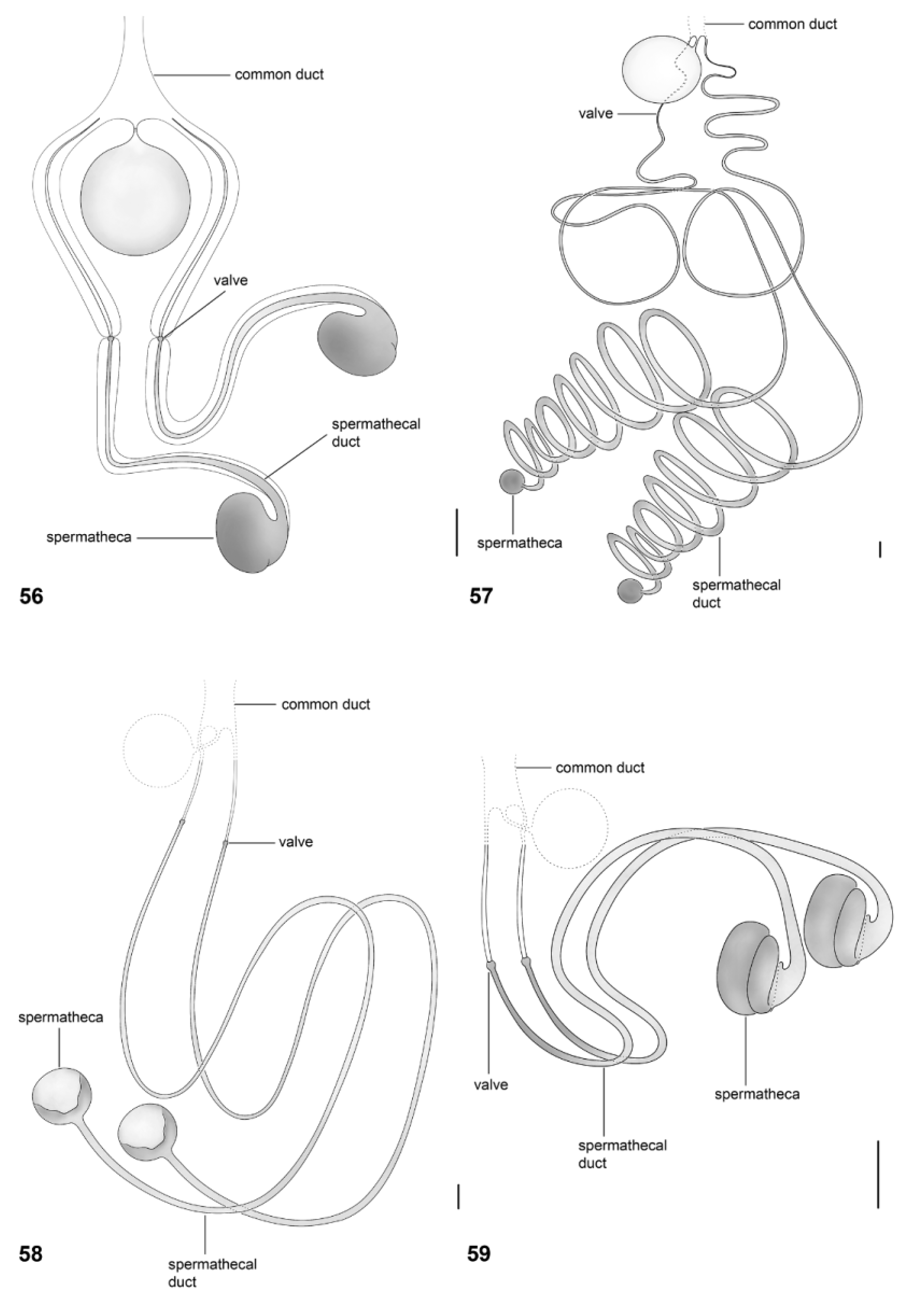
Papavero & Artigas (1991) proposed that the spermathecal capsule formed by three “layers” or coils—based only on their examination of Arthropeina fulva —is apormophic in species of the genus. This condition is actually shown here to be autapomorphic for A. pseudofulva ( Fig. 53 View FIGURES 50 – 55 ), while A. fulva has the spermatheca formed by two coils ( Fig. 18 View FIGURES 15 – 20 ). The coiled spermathecal capsule, as described by Papavero & Artigas (1971) (independently of the number of coils), seems to be a synapomorphy for a small clade within Arthropeina including A. fulva , A. colombiana and A. pseudofulva . The “remnant of middle spermathecal duct” ( Papavero & Artigas, 1991) certainly corresponds to the spermathecal duct ( e.g., Fig. 56 View FIGURES 56 – 59 ) cut during preparation, as already stated by Woodley (2011: 422). The statement that the valves of the spermathecal ducts ( Papavero & Artigas, 1991) are an apomorphic condition in the genus, hence, as commented by Woodley (2011), is not confirmed. Other dipteran families also have this condition (see e.g., Artigas, 1971). A wider comparison with other groups of lower brachycerans would be necessary to properly establish the level of origin of this feature—a single origin with losses or independent origins in different families. Additionally, the genital fork in Arthropeina presents a unique condition in Xylomyidae at the level of current knowledge, the membranous lobes ( e.g., Fig. 21 View FIGURES 21 – 26 ), not found in other xylomyid genera.
A careful morphological study of Solva species would certainly help to better understand of the evolution of features varying within Arthropeina and furnish important evidence towards the construction of a detailed phylogenetic study of the family.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |