
Patagosmilus goini
|
publication ID |
https://doi.org/ 10.5281/zenodo.196838 |
|
DOI |
https://doi.org/10.5281/zenodo.6200141 |
|
persistent identifier |
https://treatment.plazi.org/id/5D4B87BC-8B25-FFAC-CF87-FD3FDAA1FED5 |
|
treatment provided by |
Plazi |
|
scientific name |
Patagosmilus goini |
| status |
|
Patagosmilus goini gen. et sp. nov.
Figures 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5
Derivation of name. Goini , in honor of the tireless work and patient teaching of Dr. Francisco J. Goin, Argentine paleontologist whose focus is South American metatherians.
Holotype. MLP 07-VII-1-1, most of the left side of the skull with upper dentition, right magnum, and proximal portion of ungual phalanx.
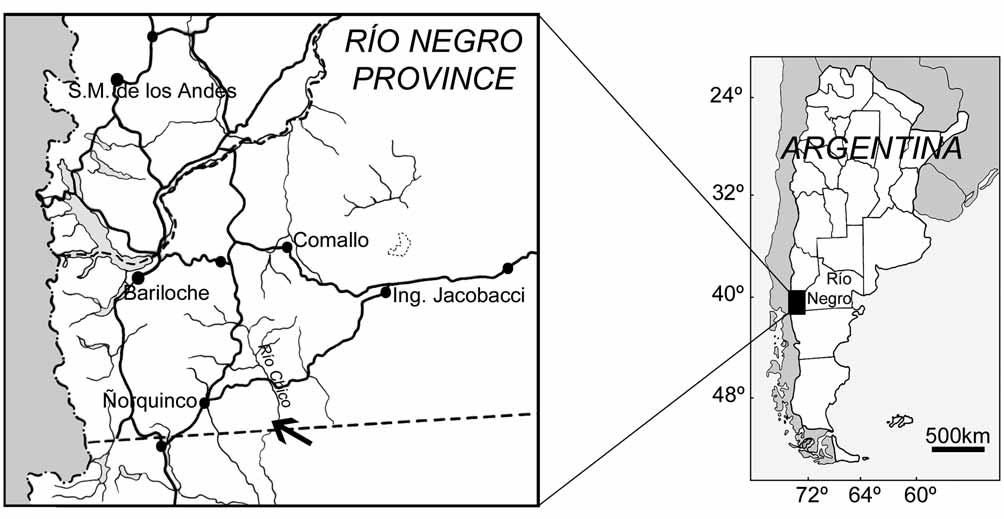
Occurrence. West margin of the Río Chico, Río Negro Province ( Fig. 1 View FIGURE 1 ), from levels of the Collón Curá Formation ( Ravazzoli & Sesana 1977), Colloncuran SALMA, middle Miocene ( Goin & Carlini 1993a, b).
Along the margin of the Río Chico, fossil-bearing exposures of the Collón Curá Formation (about 850 m. s.l.) overlay metamorphic basement, Tertiary basalts, and the Miocene La Pava Formation and are in turn overlaid by Pliocene deposits of the "Rionegrense" ( Ravazzoli & Sesana 1977; González Díaz 1990). During fieldwork, the outcrops were subdivided into northern (Duke University/ MLP Fossil Localities RCH-018 and RCH-018N) and southern exposures (RCH-018S). The specimen of Patagosmilus comes from the latter.
Diagnosis. Patagosmilus goini is smaller than Thylacosmilus and Anachlysictis . It differs from Anachlysictis in the following derived features: conspicuous bowed postcanine tooth row and short distance between the glenoid cavity (or condyle) and postmetacrista of M3 (or preprotocristid of m4). It differs from Thylacosmilus in the following plesiomorphic features: slightly longer skull, maxilla confined to the lateral side of the skull (i.e., not exposed in dorsal view), nasals widely exposed in dorsal view, presence of a subtle lacrimal process, continuous orbit and temporal fossa, lack of an anteorbital fossa and anterior notch of the orbit, longer sagittal crest, slightly shorter and straighter upper canine, P2 double rooted, molars with small protocone, M3 with narrow stylar shelf and slightly shorter postmetacrista. In addition, features unique to Patagosmilus goini are: P2 equidistantly separated from the canine and P3, sphenopalatine foramen more anteriorly located, and anterior half of nasals extremely narrow, widening conspicuously at the level of the orbits.
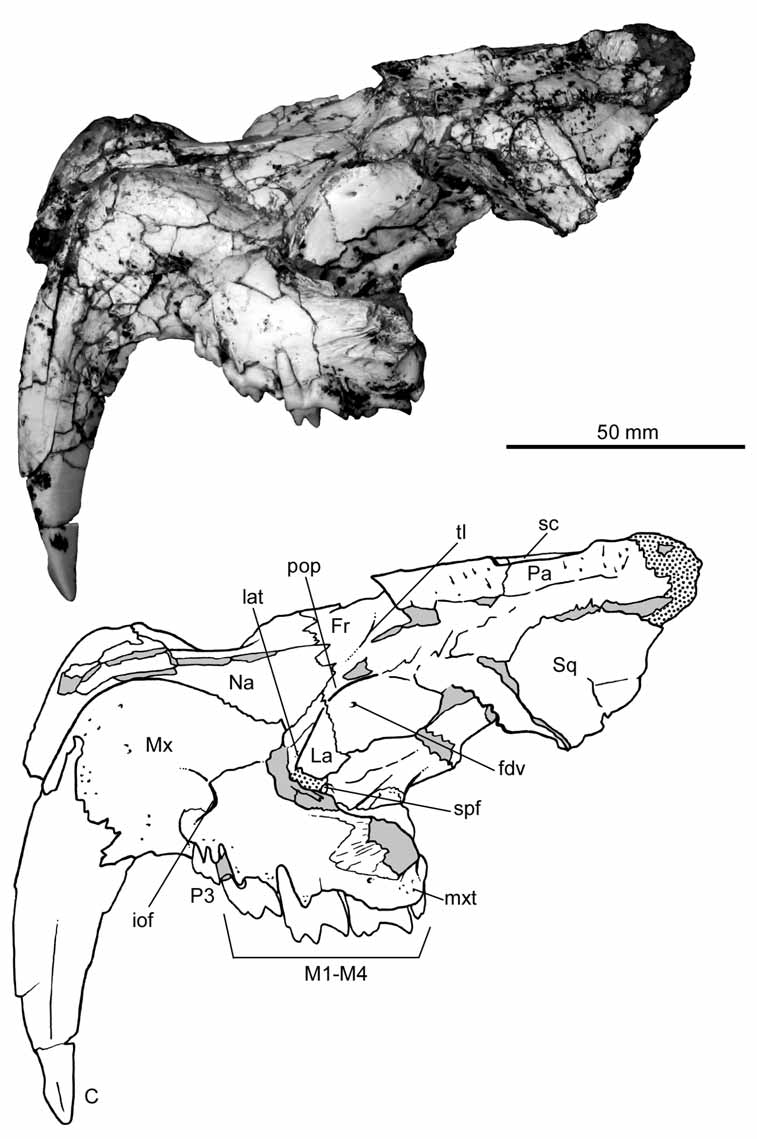
Description and comparisons. Skull. The skull of the holotype of Patagosmilus goini is partially broken and laterally compressed ( Fig. 2 View FIGURE 2 ). The left side is better preserved, although the bone is mostly fractured. The snout is short and high with two premolars and the typical hyperdeveloped upper canine that characterizes the family. The back of the skull is also high and short, though somewhat less so than in Thylacosmilus . In Patagosmilus , the nasals are widely exposed in dorsal view; very narrow anteriorly and broad posteriorly, widening at the level of the orbit ( Fig. 2 View FIGURE 2 ). This difference in width is even more conspicuous than in other sparassodonts ( Sinclair 1906; Babot et al. 2002; Forasiepi 2009). In Thylacosmilus , in contrast, the nasals are almost completely hidden in dorsal view. The nasals are apparently long, as evidenced by the section of the skull ( Riggs 1934), but developed under the maxilla; the portion seen in dorsal view is triangular, narrowing posteriorly, and restricted to the anterior part of the snout ( Riggs 1933, 1934). In Patagosmilus , the suture between nasals and frontals defines a very open “U” outline in dorsal view, as in other sparassodonts ( Sinclair 1906; Forasiepi 2009). Thylacosmilus is the exception: the frontal is not in contact with the nasal but with the maxilla, because of the enlargement of the latter (e.g., Riggs 1934; Marshall 1976; Goin & Pascual 1987; Muizon 1999). In Patagosmilus , the maxilla and the frontal are separated by the nasal and lacrimal, resembling other sparassodonts. Furthermore, the maxilla of Patagosmilus is anteroposteriorly short and high, and confined to the lateral aspect of the skull ( Fig. 2 View FIGURE 2 ). This condition differs from that of Thylacosmilus , in which the maxilla projects dorsally and posteriorly up to the level of the orbit. The root of the canine defines a prominent swelling (juga alveolaria) at the anterior border of the maxilla, which projects dorsally, following the suture with the nasal. Similar to Thylacosmilus , several minute vascular foramina open on this surface. The infraorbital foramen is relatively small, located dorsal to the posterior root of the last premolar (P3; see below); there is no anteorbital fossa, such as that found in Thylacosmilus ( Riggs 1933, 1934). At the level of the postcanine teeth, the maxilla curves laterally, defining a medially concave arcade, similar to Thylacosmilus ( Riggs 1933, 1934). In more generalized sparassodonts, in contrast, the postcanine upper tooth row is laterally straight or nearly straight ( Sinclair 1906; Babot et al. 2002; Forasiepi 2009). There is a weakly-defined maxillary tuberosity developed above the last molars. In ventral view, the lateral border of the maxilla is nearly flat with regard to the labial border of the teeth, similar to Thylacosmilus . In contrast, in several more generalized sparassodonts (e.g., Cladosictis , Arminiheringia, Callistoe , Borhyaena , Arctodictis ; Marshall 1981; Babot et al. 2002; Forasiepi 2009), the maxilla protrudes laterally from the labial border of the teeth, forming “cheeks”. In ventral view, the palate is mostly fractured; sutures between bones are not observable. The lateral border of the palate, which would correspond to part of the maxilla, is very high ( Fig. 3 View FIGURE 3 B), similar to Thylacosmilus ( Riggs 1933, 1934; Goin & Pascual 1987). There are several minute nutrient foramina on this bone surface and deep circular depressions between the molars to house the tip of the protoconid of the lower molars when the jaws are closed. The back of the palate, which would correspond to the palatine portion, is slightly thicker than the rest of the horizontal plate, forming the border of the choanae. At mid-line, the bone is broken. However, based on the fracture and the condition seen in Thylacosmilus and other sparassodonts, it is probable that there was a median nasal spine ( Sinclair 1906; Riggs 1934; Babot et al. 2002; Forasiepi 2009). The posterior part of the minor palatine foramen is preserved, seen towards the lateral side of the palate and in ventral view. This foramen was apparently large and oval. As suggested by the irregular, suture-like surface of the palatine as seen in dorsal view, the minor palatine foramen was closed at the front by a now-missing portion of the maxilla, as frequently occurs among sparassodonts ( Sinclair 1906; Riggs 1933, 1934; Babot et al. 2002; Forasiepi 2009). It is also worth mentioning that in at least one specimen of Thylacosmilus (MLP 35-X- 41-1; Riggs 1934:figure 4), the minor palatine foramen opens entirely in the palatine bone. The palatine and maxilla are exposed in the floor of the orbit. Unfortunately, however, the bones are fractured in this area and anatomical details are not discernible. At the junction of the floor and lateral wall of the orbit, a notch in the palatine bone is observed that would represent the dorsal border of the sphenopalatine foramen ( Fig. 2 View FIGURE 2 ). This aperture is close to the anterior border of the orbit, while in other sparassodonts, including Thylacosmilus , this aperture is more posterior ( Riggs 1934; Forasiepi 2009). In lateral view, the lacrimal is severely damaged ( Fig. 2 View FIGURE 2 ). The orbital portion of this bone is better developed than the facial part. In the rostrum, the lacrimal extends just beyond the orbit, with the height of the facial process more than half of its width. The lacrimal tubercle is relatively weak compared with that of Thylacosmilus ( Riggs 1934) . The lacrimal foramina could not be identified unequivocally on account of breakage. However, there is a shallow notch in the orbital process of the lacrimal bone that could lead to the now-missing lacrimal foramen. Among sparassodonts, a single lacrimal foramen located inside the orbit is the generalized condition ( Sinclair 1906; Riggs 1934) and it seems probable that the same condition occurred in Patagosmilus goini .
The zygomatic arch is broken. However, the anterior base corresponding to the maxilla is deep, suggesting that the zygoma was probably robust, similar to Thylacosmilus ( Riggs 1933, 1934). The frontal bone is partially preserved and the overall morphology resembles more generalized sparassodonts and Anachlysictis among thylacosmilids ( Goin 1997) than Thylacosmilus . The postorbital process is pronounced, but does not form a postorbital bar such as in Thylacosmilus . It is noteworthy that the latter is the only sparassodont in which the orbit is completely separated from the temporal fossa ( Riggs 1933, 1934). In Patagosmilus , there is a small foramen below the postorbital process that opens forwards, and that would correspond to the foramen for the frontal diploic vein (sensu Wible 2003). The temporal lines are weak; both contact at the mid-line of the skull and broadly diverge to the lateral side. The sagittal crest, in contrast, is better defined and sharp, tall and long, resembling other sparassodonts. In Thylacosmilus , the temporal lines are comparatively stronger; both converge backwards in the skull, describing a sigmoid line. The sagittal crest in Thylacosmilus is considerably shorter and much more robust than in Patagosmilus .
The frontals contact the parietals by means of a transverse suture visible in dorsal view ( Fig. 2 View FIGURE 2 ). The entire fronto-parietal portion of the temporal fossa displays shallow scars for attachment of the temporal musculature. These scars are deeper in Thylacosmilus . In lateral view, the frontal and parietal contact the squamous portion of the temporal, which, in turn, is the only portion of this bone that is preserved ( Fig. 2 View FIGURE 2 ).
Similar to other sparassodonts, including Thylacosmilus , the squamous portion in Patagosmilus is tall, nearly reaching the dorsal surface of the skull ( Sinclair 1906; Riggs 1934; Forasiepi 2009). The most ventral portion of the squamosal that is preserved is slightly flatter to laterally concave, suggesting that this area would correspond to the posterior base of the zygomatic process. If so, then the distance between the glenoid cavity and the last molar is very short, resembling Thylacosmilus more so than any other sparassodont ( Riggs 1933, 1934).
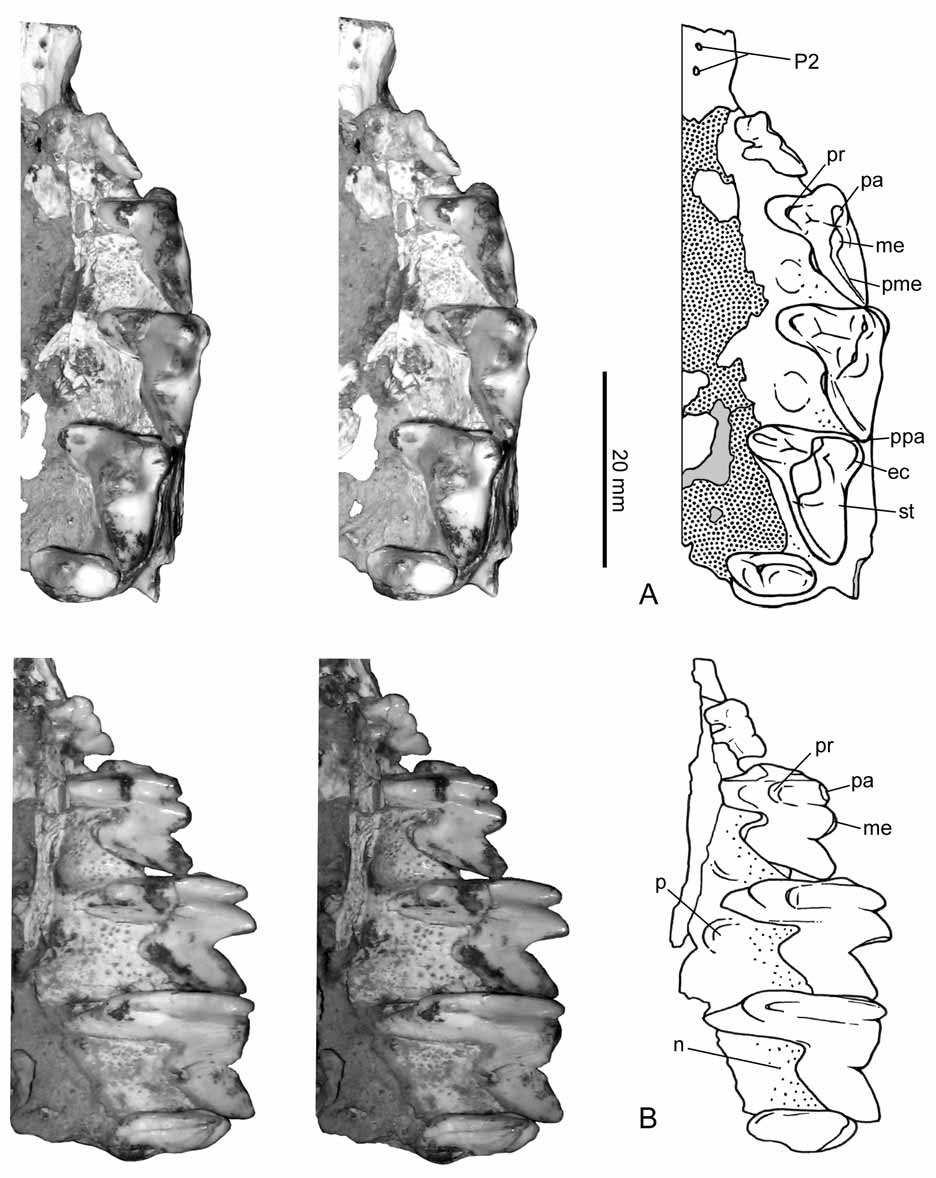
Dentition. The upper dentition is represented by one hyperdeveloped canine, one premolar (and the alveolus of a second), and four molars. This upper tooth count is also observed in Thylacosmilus and probably Anachlysictis as judged by its lower dentition ( Riggs 1933, 1934; Marshall 1976; Goin & Pascual 1987; Goin 1997). The incisors, if present, are unknown in Patagosmilus . In Thylacosmilus , upper incisors are unknown, while there are two lower incisors ( Goin & Pascual 1987). Based on wear surfaces on the lower canines, Thylacosmilus possessed at least one pair of upper incisors ( Churcher 1985). Thylacosmilus and Anachlysictis have two lower premolars. In the dentary of Thylacosmilus both premolars are similar in size and located closer to the molars than the canine. In the thylacosmilid-like taxon from La Venta (IGM 251108; Goin 1997), there are three lower premolars (the upper dentition is unknown); p1 is very small and p2-p3 are larger and similar in size. If the Colombian form in fact corresponds to a thylacosmilid, then the two lower premolars present in Thylacosmilus and Anachlysistis would correspond to p2-p3 (p1 is lost; Forasiepi 2009), and by extension, the upper premolars of Thylacosmilus and Patagosmilus would likely correspond to P2-P3 ( Fig. 3 View FIGURE 3 ).
Such as other thylacosmilids, the most outstanding feature of the upper dentition of Patagosmilus is the canine. This tooth is large, narrow, and saber-like, with a pointed tip ( Fig. 2 View FIGURE 2 ). Both anterior and posterior edges are posteriorly concave, the anterior is the most curved. The anterior border is blunt and slightly thick, whereas the posterior is sharp. In macroscopic view, the surface of the tooth is smooth, lacking grooves and crests such as those found in some sparassodonts (e.g., Arminiheringia, Callistoe , Proborhyaena , Paraborhyaena , Pharsophorus , Arctodictis , among others; Babot et al. 2002; Forasiepi 2009). The enamel is thin and restricted to the labial face of the tooth, and covers the tip and the posterior part of the blade, close to the posterior border of the tooth. The surface of the enamel has shallow wrinkles (i.e., ridges and valleys), even more conspicuous than those seen in Thylacosmilus ( Koenigswald & Goin 2000) . In lingual view, the tip of the canine of Patagosmilus has two main wear surfaces: one adjacent to the anterior border of the tooth and the other closer to the posterior one. A similar wear pattern was described for Thylacosmilus ( Goin & Pascual 1997) , suggesting a similar functional significance. The root of the canine is deeply inserted in the alveolus and it widens toward the base, suggesting that in Patagosmilus the tooth was likely hypsodont similar to Thylacosmilus , Arminiheringia, Callistoe , and probably Proborhyaena among the sparassodonts ( Simpson 1948; Marshall 1978; Babot et al. 2002; but see also Bond & Pascual 1983).
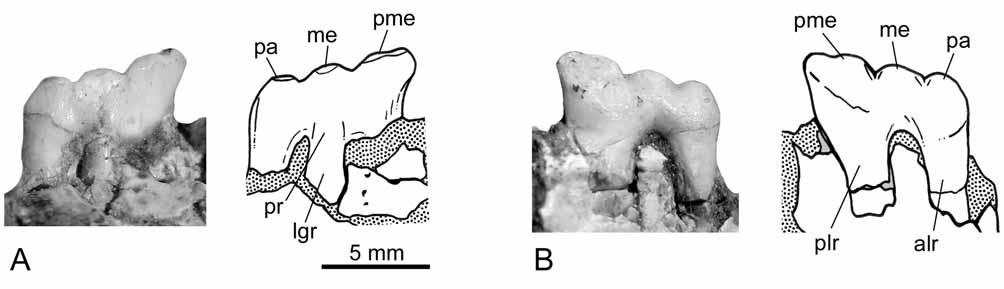
The first premolar (i.e., P2) is not preserved. There are only vestiges of what is interpreted here as the alveoli for two roots in the ventral margin of the maxilla ( Fig. 3 View FIGURE 3 ). The alveoli are minute and almost closed, suggesting that the tooth was lost in life and the bone was reabsorbed. The alveoli of P2 is set parallel to the margin of the maxilla and equidistantly separated from the canine and the following premolar (i.e., P3). In Thylacosmilus , the first upper premolar (P2) is single-rooted and set closer to other cheek teeth than the canine ( Riggs 1933, 1934; Marshall 1978; Goin & Pascual 1987). In Patagosmilus , P3 is the only premolar whose crown is known ( Fig. 4 View FIGURE 4 ). Its morphology is complex, resembling the deciduous premolar of other sparassodonts ( Sinclair 1906; Marshall 1978; Forasiepi 2009). This premolar is small compared to the molars (measurements in table 1), with three roots: one lingual and two labials, such as previously described for some specimens of Thylacosmilus ( Riggs 1933; Goin & Pascual 1997; see also MNHNA 1636, Mones & Rinderknecht 2004). The lingual root is the smallest and the posterolabial is the largest. The morphology of the crown consists of three aligned main structures, interpreted here as paracone, metacone, and postmetacrista, and a lower swelling on the lingual root that is probably homologous to the protocone. The paracone and the metacone are oval in cross section. The metacone is slightly larger than the paracone. The postmetacrista is tall and blunt. There are two notches: one between the paracone and the metacone and the other between the metacone and the postmetacrista.
The four upper molars are preserved, completely erupted in the maxilla. The wear facets are poorly defined, suggesting that specimen MLP 07-VII-1-1 is a young adult. The molars have conspicuous carnassials features, which are roughly summarized as possessing reduced crushing surfaces in favor of cutting blades ( Butler 1946; Muizon & Lange-Badré 1997). The size of the molars increases from M1 to M3, whereas M4 is small (measurements in table 1). With the exception of M4, whose morphology is simpler, the aspect of M1- M3 is largely similar among them, differing in the relative development of cusps and crests. The protocone is very narrow and considerably low, positioned at the base of the crown, while in Thylacosmilus , this cusp is absent in all upper molars and the lingual surface of the teeth is nearly vertical ( Goin & Pascual 1987). The paracone and the metacone are tall and conical. The metacone is the tallest cusp in all the molars and is more prominent in the posterior molars (except for M4). In lingual view ( Fig. 3 View FIGURE 3 B), the paracone and the metacone have two crests that descend towards the protocone. The paracone and the metacone sit very close to each other and are adjoined at their base, and for this reason the centrocrista, as such, is not identified. The postmetacrista is long and oblique in all molars, with a deep carnassial notch at the base of the metacone. In Thylacosmilus , the postmetacrista is even longer, and in M1-M3, it is half the length of the anteroposterior length of the teeth ( Goin & Pascual 1987). In Patagosmilus , there is no stylar shelf in M1; the labial face of the tooth is nearly vertical. On M2, the labial face is slightly oblique, whereas on M3, there is a narrow stylar surface. In Thylacosmilus , the stylar shelf is absent in all molars ( Goin & Pascual 1987). On M1 of Patagosmilus , there is a tiny, blunt cusp located almost at the same level as the protocone and nearly aligned with the paracone and the metacone, and this would be homologous to the parastyle. On M2, the parastyle connects with a very short cingulum that descends towards the labial side of the tooth, forming a small ectocingulum (sensu Marshall 1978). This structure is better defined in M3. The anterior cingulum is virtually absent in the molars, being only weakly present in M3. There is no preparacrista in M1-M2 and it is present only in M3, where it runs transverse to the labial border of the tooth. M4 has a simpler morphology than previous molars: it is labiolingually wide and anteroposteriorly short with only one tall cusp, the paracone. The protocone is represented by a short cingulum, whereas the metacone is absent. Differing from previous molars, the stylar region is well-developed and labial to the paracone. There is, in addition, a long and narrow anterior cingulum that descends from the anterolabial angle of the tooth to the base of the paracone. In this tooth, the preparacrista is twice larger than in M3 and nearly transversal. There is a very short postparacrista that descends from the tip of the paracone to the posterior border of the tooth.
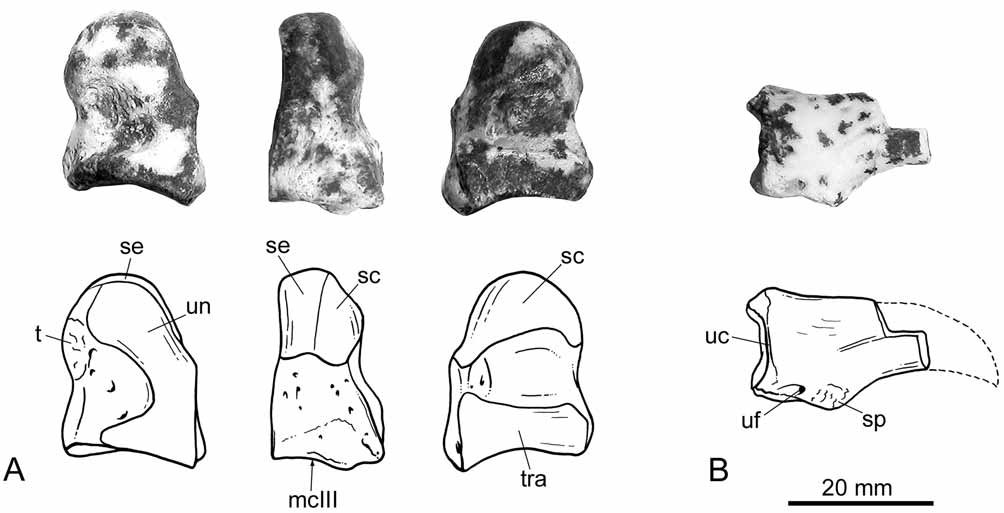
Postcranium. The right magnum and a small proximal fragment of an ungual phalanx are the only postcranial elements preserved ( Fig. 5 View FIGURE 5 ). The magnum is a small piece, very similar to the homologous element in Prothylacynus ( Sinclair 1906; Argot 2003), with a roughly rectangular outline in lateral and medial views. The lateral surface is nearly vertical and bears an elongated, articular facet adjacent to the anterior border of the bone, for articulation with the unciform. The medial surface is slightly oblique, apparently less so than in Thylacosmilus ( Riggs 1934) , and proximally convex. The proximal portion of this bone is head-like, and bears two facets: one for the semilunar in the most proximal part of the bone and one for the scaphoid, facing medially. The distal portion is massive and with straight borders. There are also two facets, one developed in medial view for articulation with the trapezoid, and the other on the distal face for articulation with the metacarpal III. The later is nearly trapezoidal in shape and slightly concave. The palmar tubercle is relatively poorly defined. Among the sparassodonts, the morphology of the magnum of Patagosmilus closely resembles that of Prothylacynus ( Sinclair 1906; Argot 2003). In Thylacosmilus , Arctodictis , and Borhyaena ( Sinclair 1906; Riggs 1934; Argot 2003, 2004; Forasiepi 2009), this bone is more pyramidal, with triangular lateral and medial surfaces, and stronger palmar tuberosities.
The ungual phalanx is very fragmentary and the preserved portion resembles that of Prothylacynus . The blade was apparently sharp dorsally, rather than blunt as in Borhyaena and Arctodictis ( Sinclair 1906; Forasiepi 2009). The articular surface is nearly circular in outline, and in this regard, it is similar to Borhyaena ( Argot 2003) . In Thylacosmilus , the articular surface is wider than tall, while in Prothylacynus , it is higher than wide ( Argot 2003, 2004). The flexor and extensor processes are subtly defined, the subungual process is small, and the ungual foramen is minute. In Thylacosmilus , in turn, the processes are stronger than in Patagosmilus .
After a broad analysis of the postcranial skeletons of several sparassodonts, Argot (2003, 2004) suggested that among large-sized forms, Prothylacynus was the most arboreally adapted taxon, whereas Borhyaena and Thylacosmilus were the most terrestrial. Because of close resemblances in the elements of the postcranium between Patagosmilus and Prothylacynus , and if the morphology of the magnum and ungual phalanx in fact reflects functional capabilities, then it is probable that Patagosmilus was able to climb.
| MLP |
Museo de La Plata |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
InfraClass |
Metatheria |
|
Order |
|
|
Family |
|
|
Genus |