
Pheretima concepcionensis, Aspe, Nonillon M. & James, Samuel W., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3881.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:FE9048E9-DE3A-4502-A95E-27EE8F706AC3 |
|
DOI |
https://doi.org/10.5281/zenodo.5670440 |
|
persistent identifier |
https://treatment.plazi.org/id/5B458787-FF9E-FFB2-FF5A-FF4CE18CBC7F |
|
treatment provided by |
Plazi |
|
scientific name |
Pheretima concepcionensis |
| status |
sp. nov. |
Pheretima concepcionensis n. sp.
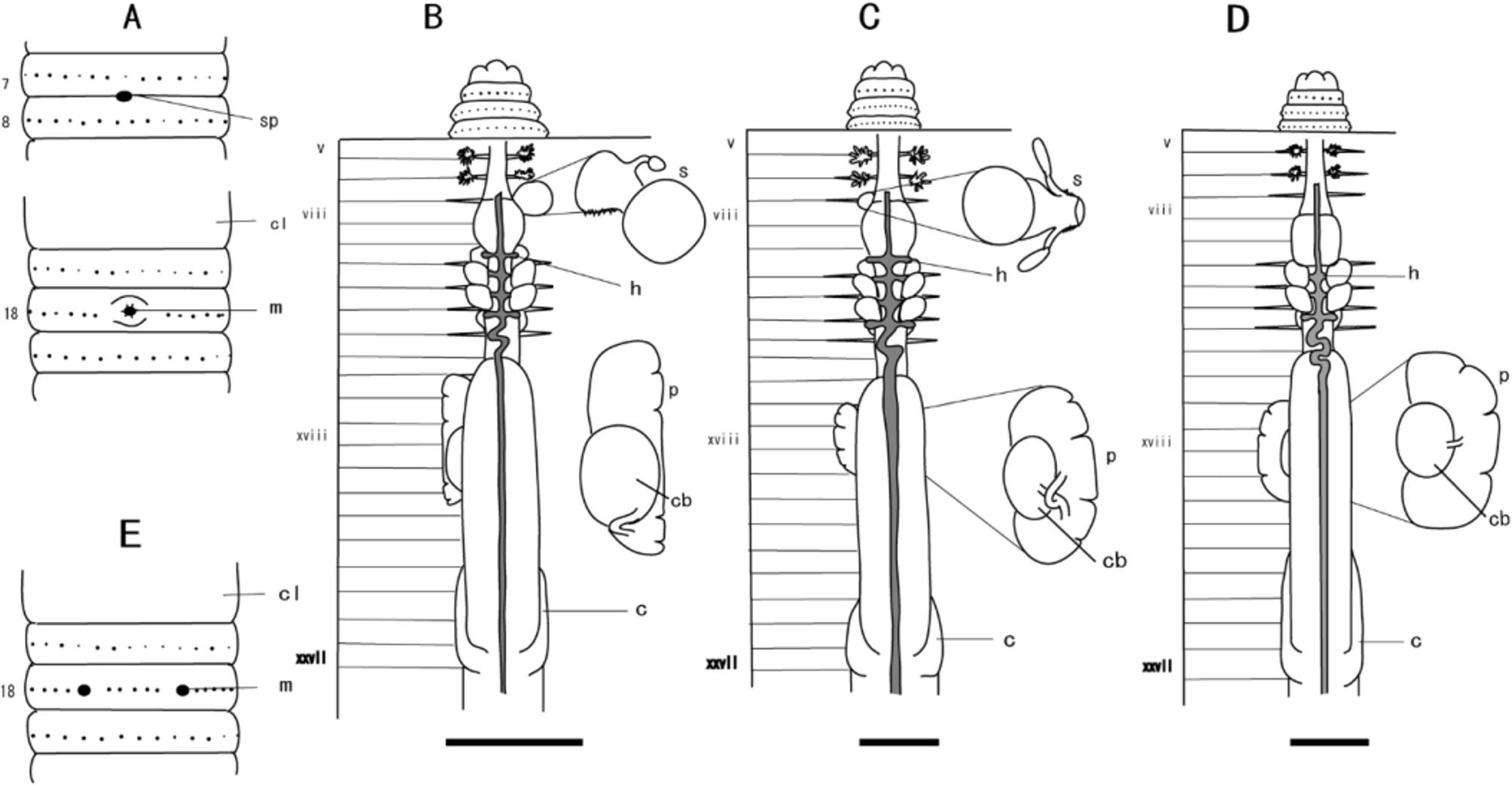
( Fig. 8 View FIGURE 8 C, Table 4)
Material examined. Holotype: adult, amputee (NMA 4522), Brgy Small Potongan, municipality of Concepcion, Misamis Occidental Province, Mt. Malindang Range (8º24'04"N, 123º36'47"E), 900 m asl., Mindanao Island, Philippines, coll. Nonillon Aspe, M. Lluch, and J. Adeva, Feb. 18–25, 2004. Paratype: one adult, posterior end missing (NMA 4542), same collection data as for holotype.
Etymology. The species name refers to the municipality of Concepcion, the type locality.
Diagnosis. Brown worm; single spermathecal pore located midventrally at 7/8; first dorsal pore 12/13; single spermatheca post-septal in viii, with nephridia on ducts; spermatheca with blocky, ovate ampulla, bulbous muscular duct expanding ectally with two stalked diverticula attached to middle of duct; prostate glands in xvii to xix; copulatory bursae confined to xviii; penis present; caeca extending from xxvii to xxv.
Description. In living animals, dorsum very dark brown anteriorly, fading to medium brown posteriorly; equators pigmented; clitellum lighter, ventral side non-pigmented. Length of holotype> 89 mm; diameter 5.2 mm at x; 4.2 mm at xx; body cylindrical in cross-section, tail narrowing gradually to sharp point;>104 segments. First dorsal pore at 12/13; single midventral spermathecal pore with elevated white lips at 7/8; female pore single in xiv; openings of copulatory bursae paired on xviii, 0.14 circumference apart ventrally, 2 setae between openings. Clitellum annular, extending from xiv to xvi. Setae evenly distributed, 40–50 setae on vii, 56–68 setae on xx, dorsal and ventral gaps present.
Septa 5/6/7/8 and 10/11–13/14 slightly muscular, 8/9/10 absent. Dense tufts of nephridia on anterior faces of 5/ 6 and 6/7; nephridia of intestinal segments located mainly on body wall at anterior and posterior faces of septa, at septum/body wall junction. Large gizzard extends from viii to x; esophagus with low vertical lamellae extends from x to xiii; intestine originates in xvi; caeca originate in xxvii, extend forward to xxv; typhlosole a simple fold of about 1/4 lumen diameter, originating at 26/27; intestinal wall with 30–35 longitudinal blood vessels.
Hearts in x to xiii, esophageal; commissural vessels in vi, vii, and ix, lateral; those in viii extend to gizzard; supra-esophageal vessel extends from x to xv; extra-esophageal vessels join ventral esophageal wall in x, receive efferent parieto-esophageal vessels in xiii.
Ovaries and funnels free in xiii; single spermatheca post-septal in viii, with nephridia on ducts; spermatheca with blocky, ovate ampulla; three spermatophores present; bulbous muscular duct expanding ectally, two stalked diverticula attached to middle of duct, terminating in ovate receptacles; stalks about same length as spermathecal duct and receptacles. Male sexual system holandric; testes and funnels enclosed in paired sacs in x and xi; seminal vesicles in xi and xii, each with long, slender dorsal lobe; vasa deferentia slender, free from body wall, passing over anterior lateral face of copulatory bursae en route to ental end of prostatic ducts; prostates in xvii to xix, each racemose, 3- to 4-lobed, wrapped around dorsal-lateral margin of copulatory bursa; muscular duct enters posterior face of copulatory bursa. Copulatory bursae in xviii hemispherical. Coelomic surface of bursae muscular, secretory diverticula lacking; floor of bursae thick, cleft by 5 furrows converging on opening; roof with blunt ovate penis.
Remarks. Pheretima concepcionensis n. sp. is similar to P. ambonensis Cognetti, 1913 and P. monoporata James, 2004 in having the spermathecal pore located in intersegment 7/8 and the spermatheca with two identical diverticula. It differs from P. ambonensis in the location of the first dorsal pore (11/ 12 in P. ambonensis ), in the presence of dorsal and ventral setal gaps (both lacking in P. ambonensis ), in the male pore spacing (0.14 circumference apart ventrally, compared to 0.2 circumference in P. ambonensis ) (James, 2004), in having the intestinal origin in xvi rather than xv; in having the spermatheca post-septal in viii (in vii in P. ambonensis ), and in the two diverticula terminating in ovate receptacles with long stalks about the same length as the spermathecal duct and receptacle (the two diverticula have much shorter chambers and stalks in P. ambonensis Cognetti, 1913 ). Pheretima concepcionensis differs from P. monoporata in size ( P. monoporata is smaller), in the number of setae (fewer in P. monoporata ), in the ventral setal gap (lacking in P. monoporata ), in septal arrangement (setae in 8/9/10 present in P. monoporata ), in the origin of the intestine (vii in P. monoporata ), in the size of the prostate (slightly larger in P. monoporata ), and in the length of caeca (longer in P. monoporata ) ( Table 4). Pheretima concepcionensis is similar to P. monotheca in having two diverticula on the spermathecal duct, but the latter species is smaller, has fewer setae, lacks ventral gaps, has the spermathecal pore in 5/6, has closer male pore spacing, has the origin of the intestine in xviii, and has longer caeca ( Table 4). Pheretima concepcionensis is similar to P. vergrandis n. sp. in the arrangement of septa, origin of the gizzard in viii, origin of the intestine, and the presence of penes, but the former species is larger, has the first dorsal pore at 12/13, has two male pores on xviii, and has shorter caeca and prostate glands ( Table 4).
Occurrence. Pheretima concepcionensis was uncommon, comprising 3.4% of individuals on plots; we found it in disturbed forest in Brgys Sibucal, Small Potongan, and Lake Duminagat, at elevations of 915–1662 m. It occurred both in the soil and above ground, on substrates such as on rotten logs (Table 1).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Oligochaeta |
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Pheretima |