
Asaphellus Callaway, 1877
|
publication ID |
https://doi.org/10.5252/g2015n2a1 |
|
publication LSID |
urn:lsid:zoobank.org:pub:85E655C9-759B-4A94-8FFB-43AE11567C1A |
|
persistent identifier |
https://treatment.plazi.org/id/480E87D9-FF9A-EC12-CAB4-FAA00B6CDDFB |
|
treatment provided by |
Felipe |
|
scientific name |
Asaphellus Callaway, 1877 |
| status |
|
Genus Asaphellus Callaway, 1877 View in CoL
Asaphelloides Kobayashi, 1937: 497.
Hemigyraspis Raymond, 1910: 41 .
Megalaspidella Kobayashi, 1937: 499 View in CoL , 500.
Plesiomegalaspis Thoral, 1946 : unnamed pages.
TYPE SPECIES. — Asaphus homfrayi Salter, 1866 , by original designation, Lower Ordovician, upper Tremadocian Stage of Porthmadog, Gwynedd, North Wales, UK.
REMARKS
Asaphids’ taxonomy is a problematic field into systematic, mainly because effacement is regarded as one of the most polyphyletic characters among trilobites ( Fortey & Shergold 1984; Turvey 2007). In addition, there are many different isoteline taxa in which effacement appears as a morphological trend (e.g., Jaanusson 1959; Nielsen 1995; Turvey 2007, among others).
It is important to note that, as Asaphellus has become a “wastebasket” taxon for the Early Ordovician (mostly Tremadocian) asaphids, an excessive expansion of its diagnosis exacerbates the problem even more ( Fortey & Owens 1987). However, some species indicate a wide range of intrageneric variation. Among them, A. whittardi ( Bates, 1969) from South Wales ( Fortey & Owens 1987) exhibits a remarkably prominently segmented pygidium for the genus, with 10 well-defined axial rings, 8 pairs of ribs, and visible interpleural furrows. Besides, its glabella is bounded by shallow but well impressed axial furrows. At the other extreme of morphological variability within the genus, in such species as A. yanheensis Yin in Yin & Shanji, 1978 (see Peng 1990) and A. isabelae n. sp., whose segmentation is nearly completely obscured by effacement, with the preglabellar furrow as an exception, because of it remains visible as a shallow furrow or a change in slope. Between these extremes, several species show very lightly impressed but discernible furrows, as in A. stenorhachis , or a diverse effacement, either of the cranidium or the pygidium. In this latter respect, A. cf. A. trinodosus Chang, 1949 from the Tremadocian of Tasmania ( Jell & Stait 1985), has an effaced cranidium but a distinct pygidial axis bounded by well-defined axial furrows. In addition, there are other cases contributing to the variability of the genus. For instance, usually the pygidial axis decreases unevenly in width and tapers backwards, however, A. lugneensis Courtessole et al., 1985 and A. fezouataensis Vidal, 1998 are examples where the pygidial axis decreases in width evenly.
Asaphellus acutulus Zhou & Zhang, 1978 from China has the entire hypostoma covered with terrace ridges (see Zhou & Zhang 1985: pl. 10, figs 4, 5), a feature that is also present in A. fezouataensis ( Vidal 1998: pl. 1, figs 2, 3), A. tataensis Vidal, 1998 ( Vidal 1998: pl. 5, fig. 2), and A. inflatus Lu, 1959 (see Peng 1990: pl. 6, fig. 8). Thus, although a terraced lateral border of hypostoma is common for the genus, a hypostoma fully covered by terraced ridges seems also a relatively frequent feature. Another consideration concerns the inner margin of pygidial border, which is not subparallel to the margin in A. stenorhachis View in CoL , which condition is not common within the genus. Finally, there are examples of anomalous features as for instance in A. stubbsi Fortey, 2009 from the Lower Fezouata Formation, where the genal spines curve outwards at genal angles before curving backwards distally, the anterior facial suture meets the anterior border without forming an ogive, and it shows rounded, forward-facing pleural extremities in its thorax. Thus, despite the apparent lack of characters of Asaphellus View in CoL due to variable effacement, there is still a wide range of morphological variety.
Some species of Asaphellus View in CoL have been erected upon poorly preserved material or upon juvenile specimens which are in need of revision. Among them, A. nanchuanensis Lu, 1975 is based upon an incomplete cranidium, a juvenile pygidium and a hypostoma. The same applies to A. inflatus longus Lu, 1975 , which was based upon very small juvenile material. Hutchison & Ingham (1967) defined Asaphoon pithogastron upon very small specimens (about 4 mm maximum widths of both pygidium and cranidium). Later, Owens et al. (1982: pl. 2, fig. g) and Fortey & Owens (1991) suggested that this material should be a young Asaphellus View in CoL meraspid. However, we suggest that the meraspid material figured in Owens et al. (1982: pl. 2, fig. g) and Hutchison & Ingham (1967: pl. 8, figs 11, 13), which includes the holotype, does not belong to Asaphellus View in CoL merapid. Only the pygidium of Hutchison & Ingham (1967: pl. 8, fig. 14) may be considered as a meraspid or an early holaspid stage of Asaphellus View in CoL . The specimens of Hutchison & Ingham (1967: pl. 8, fig. 13) and Owens et al. (1982: pl. 2, fig. g), differ mainly in the pygidial border. On the other hand, the pygidium shown in Hutchison & Ingham (1967: pl. 8, fig. 13) differs from the specimen illustrated by them (1967: pl. 8, fig. 14) in several respects, and likely they do not belong to the same species. Moreover, Asaphoon pithogastron might belong to a genus different from Asaphellus View in CoL .
In A. fezouataensis and Asaphellus cf. fezouataensis Vidal, 1998 ; the pygidial border is almost indistinct, just slightly indicated by a change in slope, whereas the cephalon exhibits a well-defined border. Moreover, A. inflatus (see Lu 1975: pl. 14, fig. 17, and Peng 1990) exhibits no border, therefore, in all respects, this species seems closer to Paramegalaspis Thoral, 1935 (Jaanusson 1956; Peng 1990) or Notopeltis Harrington & Leanza, 1957 View in CoL rather than Asaphellus View in CoL .
Although Asaphellus View in CoL includes a wide morphological variety, some assigned species do not comply with the general features of the generic diagnosis ( Fortey & Owens 1987): for instance, A. coreanicus Kobayashi, 1934 (see Choi et al. 2003) is closer to Hunnebergia Tjernvik, 1956 ; whereas Asaphellus tomkolensis Kobayashi, 1934 in Choi et al. (2003), A. communis Robison & Pantoja-Alor, 1968 View in CoL , and A. pricensis Legg, 1976 are closer to Golasaphus Shergold, 1971 . Other species such as Asaphellus sp. ( Courtessole et al. 1985), and A. cuervoae Corbacho & Lopez-Soriano, 2012 , also do not match the diagnosis of Asaphellus ( Fortey & Owens 1987) View in CoL . In addition, A. toledanus Rabano, 1990 (see also Pillet 1992) resembles Pseudomegalaspis Jaanusson, 1953 View in CoL mainly in the cranidial and pygidial doublure (see Rabano 1990: fig. 4). However, the border of the cephalon and pygidium, and the hypostoma features of A. toledanus resemble Asaphellus View in CoL . Hence, the generic status of this species requires a revision.
Asaphellus catamarcensis Kobayashi, 1935 View in CoL ( Fig. 6 View FIG )
Asaphellus View in CoL ? catamarcensis Kobayashi, 1935: 65 View in CoL , pl. 11, figs 11-15; non Asaphellus jujuanus Harrington, 1937: 115 View in CoL ; non Asaphellus ( Asaphelloides) americanus Kobayashi, 1937: 14 ; non Kobayashi, 1937a: 497; non Asaphellus catamarcensis Kobayashi View in CoL in Harrington, 1938: 246; non Harrington & Leanza, 1957: 147-151; non Tortello & Rao, 2000: 72; non Tortello, Esteban & Aceñolaza, 2002: 137; non Aceñolaza, Aráoz, Vergel, Tortello & Nieva, 2003: 23-28; non Waisfeld & Vaccari, 2003: 319; non Tortello & Aceñolaza, 2010: 162.
LECTOTYPE ( HERE DESIGNATED). — One pygidium without precise stratigraphic horizon, from Angosto de Pascha, Pascha-Incamayo area , Salta Province, USNM-PAL94136 D ( Fig. 6A View FIG ).
MATERIAL. — The type material illustrated by Kobayashi (1935: pl. 9, figs 11-15) housed in the Smithsonian Museum ( USNM- PAL 94136) comprises one cranidium, one free cheek, and two pygidia. This is refigured herein, except for the cranidium, which is lost, all from the type locality.
REMARKS
A. catamarcensis View in CoL is a much-quoted Asaphellus species from the Argentine Cordillera Oriental that has been apparently recognized in several different localities, and employed either in local or international comparisons. However, this species constitutes a remarkable nomenclatural problem and is in need of revision. It was erected by Kobayashi (1935) and placed with doubt within Asaphellus View in CoL , no diagnosis and description but only general comparisons were provided. So far, the main problems are the incomplete and poorly preserved type material, and the range of specimens showing a wide spectrum of variability that have been assigned to A. catamarcensis View in CoL since 1935 (e.g., Harrington & Leanza 1957; Tortello & Rao 2000; Tortello et al. 2002; Waisfeld & Vaccari 2003; Tortello & Aceñolaza 2010). In addition, this scenario becomes even more complex when the stratigraphic unclearness of the type locality is regarded.
The stratigraphic provenance of A. catamarcensis View in CoL is uncertain. Kobayashi (1935) explicitly noted that he received the fossils for study without locality details, except for the label (Praire Catamarca, i.e. Angosto de Pascha). According to Kobayashi (1935: 60), these fossils, preserved in a “light brown quartzite”, included Kainella meridionalis Kobayashi, 1935 View in CoL ; Kainella conica Kobayashi, 1935 View in CoL ; Kainella lata Kobayashi, 1935 ; Leptoplastides argentinensis ( Kobayashi 1935) , and Asaphellus catamarcensis ( Kobayashi 1935) View in CoL . Since Kobayashi’s work, A. catamarcensis View in CoL has been considered a common member of the “ Kainella Fauna View in CoL ”. Waisfeld & Vaccari (2003) and Vaccari et al. (2010) revised K. meridionalis View in CoL and reinterpreted the definition of the “ Kainella Fauna View in CoL ”. According to these revisions, K. meridionalis View in CoL is restricted to some boulders of the Cardonal Formation, whereas the “ Kainella Fauna View in CoL ” may occur either in the reworked boulders within debris flow deposits of the Cardonal and the Devendeus formations, or in the lower heterolithic levels of the Saladillo Formation.
From the reworked boulders of the Cardonal Formation, Waisfeld & Vaccari (2003) and Vaccari et al. (2010) reported at least two of the species listed by Kobayashi (1935): Kainella meridionalis and “ Kainella ” conica . Hence, it is highly probable that A. catamarcensis comes also from the boulders included in this unit. Alternatively, and less likely, A. catamarcensis might come from the debris flow deposits of the Devendeus Formation. The record of another Kainella species ( K. cf. K. teiichii ) in a boulder of coarse quartzitic sandstone of the Devendeus Formation (cf. Vaccari & Waisfeld 2010: figs 9.1-9.7) suggests that the provenance of A. catamarcensis from this unit cannot be ruled out.
Unfortunately the holotype cranidium figured by Kobayashi (1935: pl. 11, fig. 11) is lost, hence, a possible comparison of this sclerite is restricted to Kobayashi’s drawing. We reillustrate here a free cheek and two pygidia from the type series. The material is fragmentary and poorly preserved. The free cheek is subtriangular and the genal spine is broken off, however, a medium size might be inferred considering the width of the base. Within the type material, we are uncertain whether the two pygidia are conspecific, and because one of them is a juvenile, we have selected the specimen illustrated in Fig. 6A View FIG herein as the lectotype of A. catamarcensis . The latter is semicircular, convex, with a well-marked, anteriorly wide (tr.), and slightly funnel-shaped axis. The pygidium has a remarkably wide border (sag., exag., and tr.), being among the widest known in the genus; the doublure is wider than the border and strongly terraced.
Harrington (1938) and Harrington & Leanza (1957) redescribed the species, and gave a diagnosis, but expanding the concept of A. catamarcensis including a wide array of features exhibited by different morphotypes from more than ten localities (from Salta, Jujuy, and La Rioja provinces) ranging in age from the Furongian to the late Tremadocian (cf. Waisfeld & Vaccari 2003). Also Hoel (1999) mentioned two morphological variants, one younger than the other, both assigned to A. catamarcensis among the illustrations provided by Harrington & Leanza (1957). The incomplete nature of some material largely prevents accurate comparisons and assignments, however, the shape and definition of the axis, the effacement of the axial rings and pleural furrows, and the wide border of the pygidium selected as the lectotypepygidium suggest that none of the specimens illustrated by these authors can be considered to be conspecific with A. catamarcensis .
Harrington (1938: pl. 13, figs 3-6, 8-10, 16) referred several specimens from different localities to A. catamarcensis : the material illustrated in his figures 3 and 8 comes from the Quebrada de Iruya and it is possibly associated with K. meridionalis (according to the revision by Vaccari & Waisfeld 2010). The pygidium in Figure 3 View FIG resembles A. stenorhachis here defined, mainly in the indistinct segmentation and morphology of the border. The latter characters distinguish it from those of the lectotype of A. catamarcensis . The hipostomes ( Harrington 1938: pl. 13, figs 4, 6) are from Angosto de Pascha, the type locality of A. catamarcensis , the one illustrated in Fig. 6 View FIG does not appear to belong to an Asaphellus species. The rest of the pictures assigned to A. catamarcensis in Harrington (1938: pl. 13) match with new species under study from Quebrada de Humacha and also some of them were discuss recently in Tortello & Esteban (2014).
Megalaspis americana ( Asaphellus ( Asaphelloides) americanus Hoek in Steinman & Hoek, 1912) sensu Kobayashi (1937) is a poorly known species from Cuesta de Erquis ( Tarija Department, Bolivia). Harrington (1938: pl. 13, figs 19, 20) assigned two distorted pygidia from Río Volcancito (Sierra de Famatina, La Rioja) to Asaphellus ( Asaphelloides) americanus and Harrington & Leanza (1957: figs 65.3, 5) referred the same specimens to A. catamarcensis View in CoL considering that the Bolivian species and A. catamarcensis View in CoL were synonymous. Among other features, the well-impressed segmentation of the pleural fields suggests that the material from Río Volcancito is not conspecific with A. catamarcensis View in CoL , whereas the validity of Megalaspis americana requires further corroboration.
The material illustrated by Harrington & Leanza (1957: figs 65.7, 8) from Quebrada de Los Canchos (Caldera Department, Salta) was the basis for the concept of A. catamarcensis View in CoL of Tortello & Rao (2000), Tortello et al. (2002), Aceñolaza et al. (2003), and Tortello & Aceñolaza (2010). The outline, shape of the axis and the border, and segmentation of the pygidia do not conform to the lectotype of A. catamarcensis View in CoL . Instead, this material is referred here to Asaphellus sp. 1 ( Fig. 7 View FIG C-F) from the Quebrada del Arenal, Rupasca Member, Kainella teiichii Zone (Vaccari et al. 2010) . In addition, Tortello & Aceñolaza (2010) also referred to A. catamarcensis View in CoL two cranidia that the authors considered very close to the type cranidium illustrated by Kobayashi (1935). The cranidia from Abra de Zenta ( Tortello & Aceñolaza 2010; Fig. 4A, B View FIG ) shows features such as a strongly tapering glabella with a pronounced lateral constriction at midlength (exsag.), and a quite deeply impressed preglabellar furrow. Despite the fragmentary nature of the cranidium figured by Kobayashi (1935) that prevents detailed comparisons, neither the lateral constriction nor the deeply impressed preglabellar furrow are visible. Hence, we do not agree with the assignment of these sclerites to A. catamarcensis View in CoL . In addition, the pygidium in Harrington (1938: pl. 13, fig. 10) from Angostura de Parcha is identical to that of Asaphellus sp. 1 reported herein.
Several of the specimens referred to A. catamarcensis by Harrington & Leanza (1957: figs 65.2, 4, 6; 64.4, 5, 6) come from the upper Tremadocian successions of the Quebrada de Coquena, Quebrada de Chalala, and Río Santa Victoria. They are apparently close to a new species that it is now under revision by the authors from the upper middle Tremadocian successions exposed in Quebrada de Humacha and Quebrada de Chalala, and also close to the recent published asaphids in Tortello & Esteban (2014).
Likewise, the pygidia from Río Volcancito ( La Rioja, Furongian, Parabolina ( N.) frequens argentina Zone, Harrington & Leanza 1957 : figs 65.3) are distinguished from that of A. catamarcensis in a fairly segmented, shorter, and wider axis. Finally, the pygidia from Quebrada Colorada (Iruya, K. meridionalis Zone ), referred to A. catamarcensis by Harrington & Leanza (1957: pl. 64.1, 65. 1), differ from the lectotype pygidium in a more well-segmented axis and narrower border.
Waisfeld &Vaccari (2003: pl. 22, figs 3-6) assigned to A. catamarcensis some material derived from a sandstone boulder of the Cardonal Formation in the Pascha-Incamayo region. This boulder comes from the alleged type locality of Kobayashi’s material and contains K. meridionalis . The pygidium figured by Waisfeld & Vaccari (2003: pl. 22, figs 4, 5) is similar to the lectotype in several respects, however, the axis is less-defined and more parallel sided; the border is incomplete but appears to be narrower than the lectotype.The pygidium of Waisfeld & Vaccari (2003: pl. 22, fig. 4, 5) is reinterpreted here as A. isabelae n. sp.
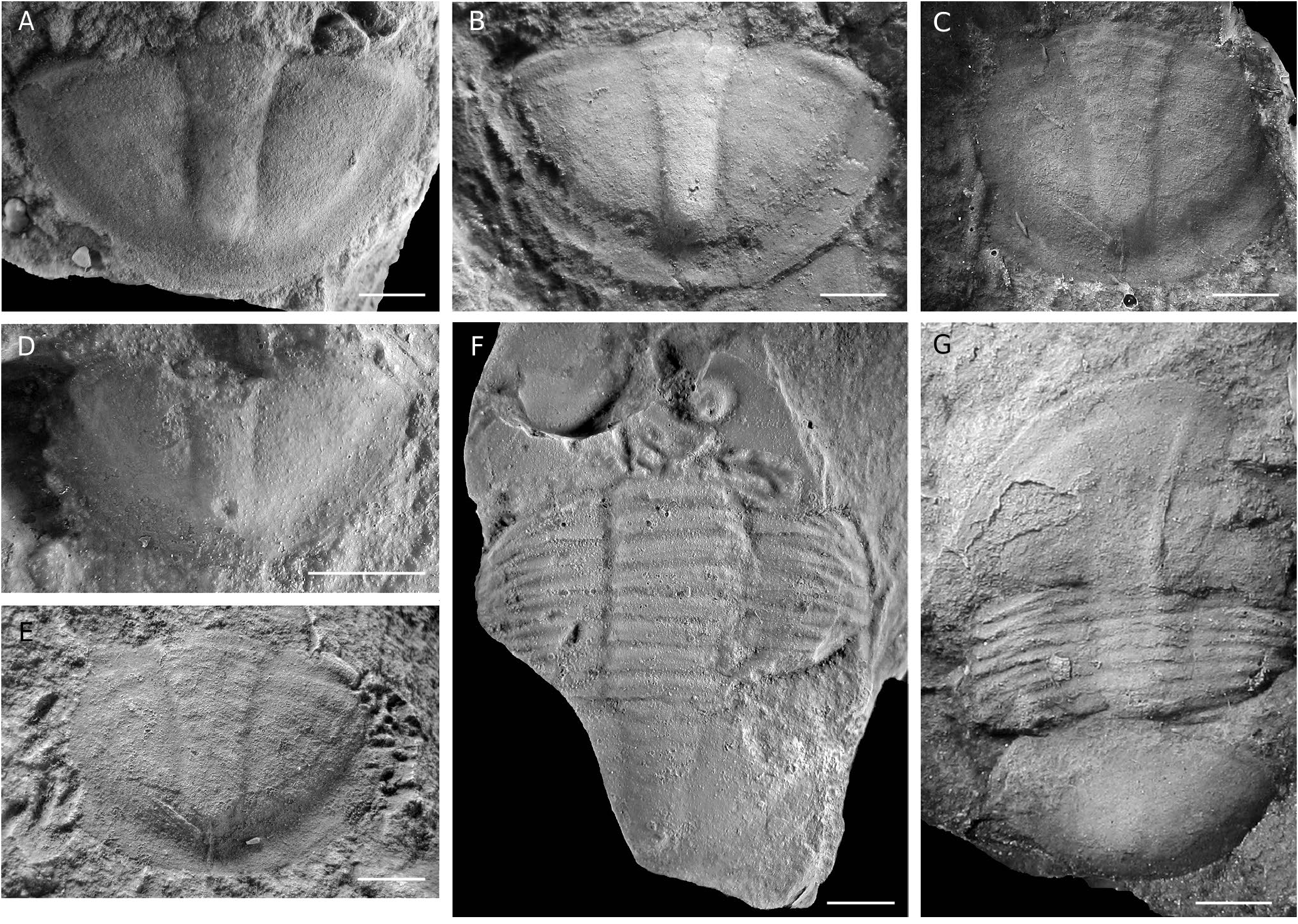
Asaphellus stenorhachis ( Harrington, 1938) View in CoL ( Figs 8 View FIG A-M, 9A)
Illaenus stenorhachis Harrington, 1938: 182 pl. 5, fig. 18.
Illaenopsis stenorhachis View in CoL – Harrington & Leanza 1957: 183, figs 93.3; non figs 93.1, 2, 4.
HOLOTYPE. — CPBA (collection of the UBA – Universidad Nacional de Buenos Aires) no. 4165 ( Harrington 1938: pl. 5, fig.18; Harrington & Leanza 1957: fig.93.3), Quebrada de Rupasca,Alfarcito region,Rupasca Member, Santa Rosita Formation, Santa Victoria Group, lower Tr2.
MATERIAL. — 57 fragmentary specimens including 3 partially complete individuals, 21 cranidia, 27 pygidia, 15 free cheeks, 8 specimens with thoracic segments, and 3 hypostomas. Illustrated material CEGH-UNC 25736-25748.
OCCURRENCE AND AGE. — Quebrada del Arenal (levels from DLA 31 to DLA 37). Quebrada Rupasca, and Angosto de Chucalezna, J. Til.- 2 in Harrington & Leanza (1957). Quebrada Colorada and Angosto de Chucalezna, S. Iru. 4 in Harrington (1938). Rupasca Member, Santa Rosita Formation, Santa Victoria Group. Middle Tremadocian (Tr2), B. tetragonalis Zone.
EMENDED DIAGNOSIS. — Exoskeleton elongated and elliptical, about 70% longer than wide. Cranidium smooth, glabella almost indistinct laterally, preglabellar furrow ill-defined, convex forward. Genal spines widely based, and short (about 20% of free cheek maximum length). Pygidial border flat to slightly concave, wider anteriorly than posteromedially, barely defined axis in the anterior third of the pygidium.
DESCRIPTION
Cephalon semicircular, slightly convex, wide and depressed frontal area without differentiated anterior border, 1.8 times wider than long and 40% of the total body length, wide (tr.) posterolateral projection. Lateral border broad and well- marked by a change in slope, posterior border defined by a wide (exsag.), shallow, and transverse posterior border furrow.
Facial suture isoteliform, anterior branch slightly divergent in front of eyes, meeting anteriorly in a tiny ogive; posterior branch sigmoidal, runs outwards and backwards behind the eyes, and slightly inwards cutting posterior border close to the genal angle.
Cranidium almost flat; palpebral, preocular, and postocular areas barely discernible. Preglabellar area flat, defined by a gentle change in slope, reaching about 15% of cranidial maximum length (sag.). Wide posterior area of fixigena (exag., tr.). Glabella smooth, defined by indistinct axial furrows, with no differential convexity compared with the rest of the cranidium ( Fig. 8A, B View FIG ); occipital ring reduced, poorly defined by a shallow furrow, slightly convex forward, set close to posterior margin. Median node located at about 1⁄5 of the cranidial maximum length from posterior margin.
Palpebral lobe of medium size, flat, broadly arcuate, located at cranidial midlength, palpebral furrow nearly effaced; eye length (exag.)/cranidial maximum length (sag.) about 0.22 (n =7).
Free cheek subtriangular, broadly arcuate external margin; doublure subparallel to the margin, lateral border defined by broad and shallow furrow, diminishing in width and fading out towards genal angle, depressed with respect to genal field, border narrow and weakly convex. Genal spine broad-based, and short (about 20% of the librigena length), strongly tapering and directed straight backwards ( Fig. 8B, E View FIG ).
Hypostoma elongate (sag.) and suboval, with rounded posterior margin; wide posterolateral borders with strong and closely spaced terrace lines. Median body gently convex, longer than wide, posterior lobe crescentic, maculae sharply impressed, transverse, and deep; anterior wings wide (tr.) and subrounded.
Thorax with 8 segments, about the same length (sag.) as the pygidium, decreasing slightly in width (tr.) backwards. Axis weakly convex, about 35% of the total width (tr.), defined by well impressed and slightly backwards-tapering axial furrows; articulating furrow narrow and deep; articulating half-ring short (tr.). Pleural region moderately wide (about 65% of thoracic maximum width); pleural furrow deeper and wider medially, runs toward anterior margin of pleura, shallower and narrower adaxially, lightly impressed abaxially, anterior pleural band slightly wider (exag.) than posterior pleural band. Pleural extremities obliquely truncated and facing backwards, well-marked pleural fulcra.
Pygidium slightly convex, about 55% wider than long. Border flat, well-defined by a shallow furrow and a change in slope, decreasing progressively in width backwards, narrowest posteromedially. Anterolateral corner of pygidium (articulating facet) subtriangular. Inner margin of doublure subparallel to the margin. Fine and closely spaced terrace lines visible on doublure (on exfoliated material). Axis gently convex (tr., sag.), about 20% of pygidial width, bounded by weak axial furrows, indistinct posteriorly ( Fig. 8E, G, K View FIG ), almost effaced in some specimens ( Fig. 8I, J, L, M View FIG ), evenly tapering backwards, posterior tip slightly raised, terminating in advance of border furrow. Articulating half-ring of even width, defined by a transverse furrow. 2-3 anterior axial rings slightly impressed in specimens preserved in mudstone, defined by narrow and shallow ring furrows, up to eight faint rings visible in some specimens (mostly internal moulds).
Pleural field wide, subtriangular; inner part gently convex (tr.), then steeply curving down to border furrow; anterior halfrib well defined by distinct furrow, wider medially, abruptly terminating at border furrow, remainder ribs and furrows faint or indistinct.
REMARKS
Harrington (1938) erected Illaenus stenorhachis on the basis of a single pygidium with four attached thoracic segments from the Quebrada Rupasca (Alfarcito area). Later, Harrington & Leanza (1957: 183, 184) included in this species additional material from other localities of the Cordillera Oriental (Quebrada Lampazar and Quebrada Pingüiyal), and accommodated it in Illaenopsis Salter, 1867 . Waisfeld & Vaccari (2003) noted that the holotype is an asaphid, and hence the name stenorhachis should be restricted to it. Although the holotype ( Harrington 1938: pl. 5, fig. 18; Harrington & Leanza 1957: fig. 93.3) is slightly distorted, the Asaphellus material from the Quebrada del Arenal studied herein closely resembles it in an identical pygidial outline, mostly indistinct axial and pleural furrows (except for the first pleural furrow), and the uneven width of the border, wider laterally than posteromedially. Hence, the material from Quebrada del Arenal is referred herein to Asaphellus stenorhachis . Asaphellus stenorhachis is similar to the type species A. homfrayi Salter, 1866 from the late Tremadocian Garth Hill and Shineton Shale formations (North Wales and Shropshire, UK) ( Lake 1942; Fortey & Owens 1991) in the general effacement of the exoskeleton and the anterior course of the facial suture with a similar variation range between the anterior and posterior branches (90-110°). However, in A. stenorhachis the glabella is less defined laterally, the preglabellar field is narrower (sag., exsag.), the postocular fixigena is wider (tr.), the genal spine is shorter, the pygidial border is of uneven width (sag., exsag., tr.) and finally, the pygidial axis is less distinct and less raised above the pleural field.
Several species of Asaphellus are known from north western Argentina. A. catamarcensis , herein restricted to the type material, distinguishes from A. stenorhachis in the pygidial outline, wider border, well-defined axis, and more segmented pleural fields.
Asaphellus jujuanus Harrington, 1937 View in CoL is a poorly known species from the upper Tremadocian of the Quebrada Coquena, Jujuy Province ( Harrington 1937, 1938; Harrington & Leanza 1957: figs 66, 6-12; Waisfeld & Vaccari 2003). It differs from A. stenorhachis View in CoL mainly in the cranidial features, such as a narrower (exsag.) and shorter (tr.) posterior fixigenal area, larger eyes, and more distinct axial furrows defining the glabella.
Asaphellus kayseri (Kobayashi, 1937) is another Argentinean species in need of revision; its age and stratigraphic position are uncertain (cf. Waisfeld & Vaccari 2003: 320). A. kayseri ( sensu Harrington & Leanza 1957: figs 75, 1-9) differs from A. stenorhachis View in CoL in a well impressed axial glabellar furrows, subparallel anterior sections of the facial suture, and a better segmented pygidial axis and pleural fields.
Finally, the general effacement of the glabella, course of the anterior facial suture, pygidial outline and pygidial furrows of A. stenorhachis View in CoL clearly distinguishes it from A. riojanus Harrington & Leanza, 1957 View in CoL from the Furongian of the Famatina Range (Volcancito Formation; Tortello & Esteban 2007), mainly in the course of the anterior suture and the general outline of the pygidium.
The pygidium referred by Harrington (1938: pl. 13, fig. 3) to A. catamarcensis from Quebrada Colorada (Iruya area) resembles that of A. stenorhachis in several aspects, such as the general outline, degree of effacement, and shape of the border. It is possibly conspecific with A. stenorhachis , but better preserved material is required in order to be certain of this assignment.
Suárez Soruco (1975: pl. 2, figs 1, 2) defined A. palquiensis from the Kainella andina Zone , lower Tremadocian (Tr1) of South Bolivia (Vaccari et al. 2010). A. palquiensis differs from A. stenorhachis in a much longer genal spines, a better defined preglabellar area, and stronger segmentation of the pygidium.
Among other non-Argentinean species of Asaphellus, Robison & Pantoja-Alor (1968) described three species from the Tiñú Formation (Tremadocian, Oaxaca, Mexico); of these, only A. artus bears some resemblance to A. stenorhachis The smoothness and posteromedian decrease in width of the pygidium of the latter, resembles the external moulds of the Mexican species (Robison & Pantoja-Alor 1968: pl. 99, figs 1-6), however, the exfoliated material of A. artus displays a well-defined and segmented pygidial axis, and distinctly furrowed pleural fields. Besides, A. artus has larger eyes, a well-defined frontal area, and larger genal spines.
Another similar species is A. cf. graffi Thoral, 1946 ( Fortey & Owens 1992: fig. 4) from the Habberley Formation (late Tremadocian, Welsh basin, UK), however, the outline of the pygidium, the well-defined axis, and a broader border clearly distinguishes it from A. stenorhachis .
A group of Tremadocian species of Asaphellus from China described as A. inflatus Lu, 1959 ; A. tungtzuensis Sheng, 1958 ; and A. yanheensis Yin in Yin & Shanji, 1978) from the Nantsinkwan Formation of the Yangtze Platform ( Peng 1990) display a very faint segmentation in the pygidium, thus resembling A. stenorhachis , however, the hypostoma of the latter has a non-terraced median body and more marked cephalic and pygidial borders defined by a change in slope.
The nature of the pygidial border of A. acutulus Zhou & Zhang, 1985 from the Tremadocian of the Jilin region, northeast China, closely resembles that of A. stenorhachis (particularly the specimens illustrated by Zhou & Zhang 1985: pl. 10, figs 7, 8), nonetheless, considering several differences in the cranidial features, the general outline of the pygidium, the axis that is not funnel-shaped and decreases steadily backwards in width, distinguishes this from A. stenorhachis .




| PAL |
Herbarium Mediterraneum Panormitanum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Asaphellus Callaway, 1877
| Arcerito, Facundo René Meroi, Waisfeld, Beatriz & Balseiro, Diego 2015 |
Illaenopsis stenorhachis
| HARRINGTON H. J. & LEANZA A. F. 1957: 183 |
Illaenus stenorhachis
| HARRINGTON H. J. 1938: 182 |
Asaphellus
| TORTELLO M. F. & ACENOLAZA G. F. 2010: 162 |
| ACENOLAZA G. F. & ARAOZ L. & VERGEL M. M. & TORTELLO M. F. & NIEVA S. M. 2003: 23 |
| WAISFELD B. G. & VACCARI N. E. 2003: 319 |
| TORTELLO M. F. & ESTEBAN S. B. & ACENOLAZA G. F. 2002: 137 |
| TORTELLO M. F. & RAO R. I. 2000: 72 |
| HARRINGTON H. J. & LEANZA A. F. 1957: 147 |
| HARRINGTON H. J. 1938: 246 |
| HARRINGTON H. J. 1937: 115 |
| KOBAYASHI T. 1935: 65 |
Hemigyraspis
| RAYMOND P. E. 1910: 41 |