
Integripelta acanthus, Gordon, D. P. & Rudman, W. B., 2006
|
publication ID |
https://doi.org/10.5281/zenodo.273434 |
|
DOI |
https://doi.org/10.5281/zenodo.5671574 |
|
persistent identifier |
https://treatment.plazi.org/id/455787E2-9443-FFB2-FEC8-F9A4FBBF9228 |
|
treatment provided by |
Plazi |
|
scientific name |
Integripelta acanthus |
| status |
sp. nov. |
Integripelta acanthus View in CoL n. sp.
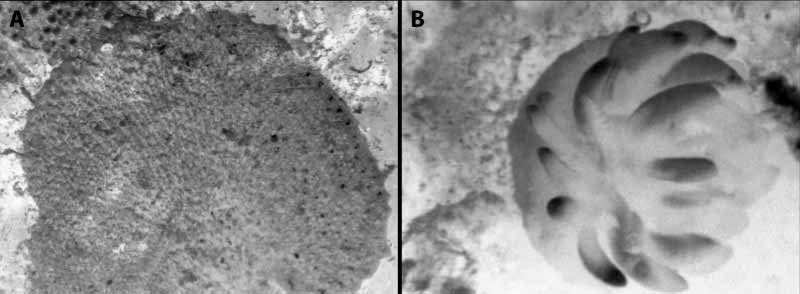
( Figures 1 View FIGURE 1 A, B)
Holotype
AM U.4706, dried, cleaned in sodium hypochlorite, encrusting on a small rock. Paratypes AM U.4723, U.4724, dried with membranes intact, on the reverse side of the same rock bearing the holotype.
Description
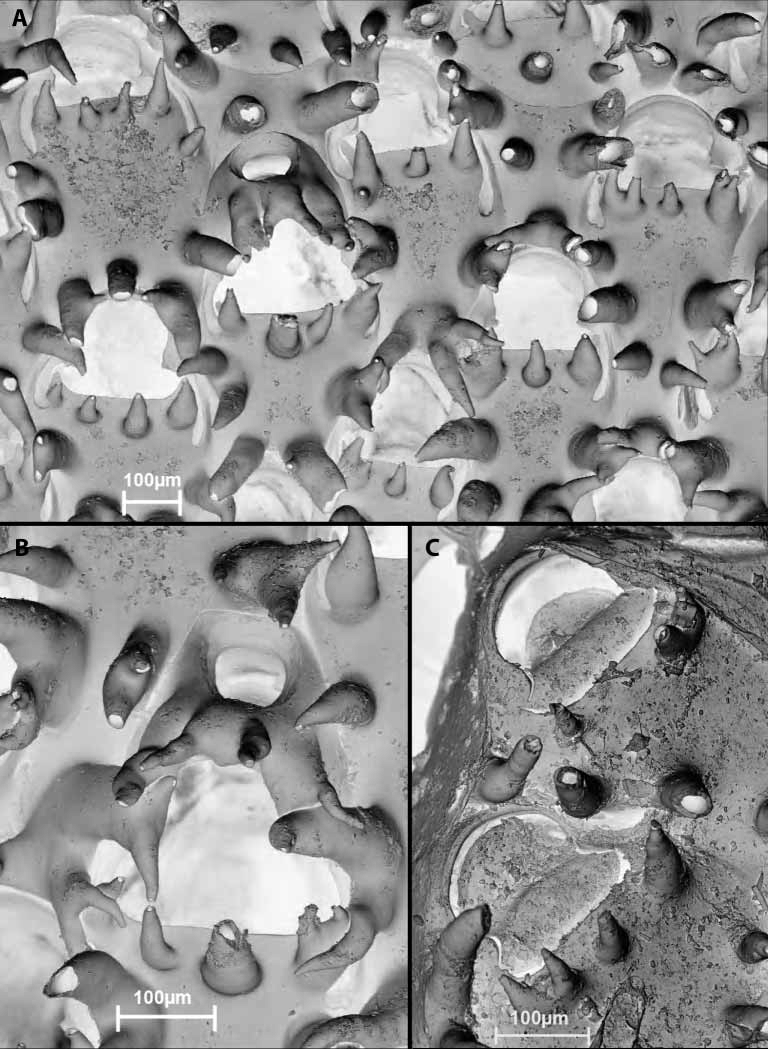
Colony encrusting, multiserial, unilaminar, attaining a size up to 17 mm diameter. Colour in life more or less pinkishred, merging to pink at colony margin. Autozooids continuous, arranged quincuncially, 380–780 µm long (mean 500 µm), 280–470 µm wide (mean 330 µm) in zone of astogenetic repetition. Gymnocystal frontal shield typically bearing stout thornlike spines, but not necessarily in all parts of the colony; where they occur there are 3–4 along the proximal rim of the orifice with additional spines around the margin that tend to be angled towards adjacent orifices; all spines are nonarticulated and hollow, some are open at the tip (not closed skeletally), and some are bifurcate. Orifice 130–170 µm high (mean 140 µm), 170–200 µm wide (mean 180 µm), with a higharched anter and welldeveloped rounded proximolateral embayments; proximal rim straight or very gently convex; the lateral excavations of the gymnocyst well developed, occurring on either side of the embayments and extending equally or unequally on each side to the vicinity of a spine or well past a spine, the sides of each excavation parallel or the excavation proximally clavate. Orifice of maternal zooids somewhat dimorphic, being larger than that of autozooids, 230–290 µm wide (mean 250 µm), with the articular condyles less angular; distal kenozooidal chamber well developed frontally with a conspicuous round foramen half its width, the chamber short, not reaching the basal wall. No basal porechambers, mural septular pores occur low in the walls of each zooid. Ancestrula 400 µm long, 270 µm wide, with a membranous frontal wall, no marginal spines.
Etymology
Latin noun ( acanthus , thorn) in apposition.
Remarks
The small rock bearing the holotype colony also bears two paratype colonies on the reverse side. The smaller of the two is an infertile colony that lacks the thornlike spines. That these spines would not have been subsequently produced by these zooids is evidenced by the larger paratype colony and the holotype colony in which a significant zone of periancestrular zooids lacks them. It is clear from the smooth frontal shield in these zooids that spines have not been broken off; they had never formed. Spines are entirely superficial; their cavity is not continuous with the zooidal chamber via holes in the gymnocystal shield. They form early in zooidal ontogeny as the gymnocystal shield is secreted, prior to complete differentiation of the zooid.
Integripelta acanthus View in CoL most resembles the Japanese species I. novella Gordon, Mawatari & Kajihara, 2002 View in CoL from Hokkaido but this species has much broader gymnocystal excavations and a smaller foramen in the ooecial kenozooid. Okada (1929) reported a spined eurystomellid from Mutsu Bay at the northern end of Honshu, Japan, that he attributed to I. bilabiata (as Lepralia View in CoL ). It has narrow crescentic slits extending proximally from each side of the orifice, like those in I. shirayamai Gordon, Mawatari & Kajihara, 2002 View in CoL , leading Gordon et al. (2002) to include Okada’s material in the synonymy of I. shirayamai View in CoL . It is now clear, from a careful comparison of Okada’s (1929) illustrations with I. shirayamai View in CoL and I. acanthus View in CoL , that Okada’s material represents an undescribed species. In addition to a median suboral spine that is continuous with a carinalike convexity of the gymnocystal frontal shield, the orifice is wider than long (longer than wide in I. shirayamai View in CoL ) and the foramen of the ooecial kenozooid is transversely oval instead of beanshaped. It is not possible to introduce a new name for Okada’s material here as the repository of his specimens is uncertain.
Ecology
The description of Integripelta acanthus above is based on several colonies on a small rock, collected from a depth of 3 m at Cape D’Aguilar Bay. Associates on the rock include other encrusting bryozoans (genera Antropora , Celleporina , Chorizopora , Electra , Fenestrulina , Hippothoa , Metroperiella , Parasmittina , Rhynchozoon ), serpulid tubeworms, foraminiferans, and crustose coralline algae.
The goniodoridid nudibranch Okenia hiroi ( Baba, 1938) has often been observed in association with I. acanthus (see Rudman & Darvell 2001; Rudman 2004a, b). Its colour matches that of the bryozoan on which it feeds ( Fig. 2 View FIGURE 2 ). Previously attributed to Hopkinsia MacFarland , O. hiroi and related species have been merged in Okenia Menke ( Gosliner 2004) . Okenia hiroi is known from the Pacific coast of Japan, Korea, and Hong Kong ( Rudman 2004b). Integripelta acanthus is so far known only from Hong Kong, but as most species of Okenia appear to be speciesspecific in their diet, it is probable that the range of I. acanthus at least includes Japan and South Korea where O. hiroi is commonly found.
Boreholes occur in several autozooids of the holotype colony of I. acanthus . They are mostly suboval, occur near the lateral margin of a zooid, and have their long axis parallel to that of the zooid. The boreholes are wider externally (100–180 µm maximum diameter) than internally (40–110 µm maximum diameter), with bevelling surrounding the inner opening. From what is known of radular morphology in Okenia hiroi , it is unlikely to have caused these boreholes (which were possibly the work of a marginelliform ( Marginellidae , Cystiscidae ) gastropod). All of the boreholes are in autozooids with spines.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Neocheilostomina |
|
Family |
|
|
Genus |
Integripelta acanthus
| Gordon, D. P. & Rudman, W. B. 2006 |
I. novella
| Gordon, Mawatari & Kajihara 2002 |
I. shirayamai
| Gordon, Mawatari & Kajihara 2002 |