
Xystopyge pelecys, Frederiksen, Sara B. & Enghoff, Henrik, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.212354 |
|
publication LSID |
lsid:zoobank.org:pub:D25AC77D-2E5B-4B99-8449-812558DB22E2 |
|
DOI |
https://doi.org/10.5281/zenodo.6172389 |
|
persistent identifier |
https://treatment.plazi.org/id/42445940-FF91-BF6F-FF21-10E1FEE4FD82 |
|
treatment provided by |
Plazi |
|
scientific name |
Xystopyge pelecys |
| status |
sp. nov. |
Xystopyge pelecys View in CoL n. sp.
Figs. 1–10 View FIGURE 1 – 7 View FIGURE 8 View FIGURE 9 – 10
Material: Holotype male: TANZANIA, Tanga region, Muheza district, Nilo forest reserve 04°54`16”S 38°39`45”E, 09/VI/2000, Frontier Tanzania, ( ZMUC 00020501). Paratypes 2 males: same data as holotype, ( ZMUC 0 0 0 20521, ZMUC 00101494). 1 male: TANZANIA, Tanga region, Muheza + Korogwe districts, Nilo Forest reserve, 04°57`33,1”S, 38°38`28,6”E. 13/II/2001, Frontier Tanzania, ( ZMUC 00020511). 1 male: TANZANIA, Tanga region, Muheza district, Nilo Forest reserve, 04°51'S, 38°39'E, 24.IX.2000, Frontier Tanzania, ( ZMUC 00020516). 2 males: Tanzania, East Usambara mts, Amani, 1000 m. 10/VII/1980 and 20/VII/1980, M. Stoltze and N. Scharff leg. ( ZMUC 00020506), ZMUC 00020505).
Diagnosis: Metaplica large, sickle-shaped, proplica very small in comparison. Differs from congeneric species by the broad flat rounded lateral metaplical process and the axe-head-shaped process on the telomere, the shape of both processes are unique.
Etymology: The name is a noun in apposition and refers to the axe-shaped telomeric process ( pelecys, Greek for axe).
Description (based on all specimens):
Body length 59–64 mm, diameter 3.5–4.0 mm. 62–65 podous body rings, no apodous rings in front of telson.
Coloration (after 11 years in alcohol): Head and antennae mostly brown, but with two lighter brown/yellow spots between the antennae and a darker band between the eyes. Supralabral region yellow, labrum brown. Collum brown with yellow edges. The rest of the body rings mostly brown (although much is faded to grey in alcohol, original colour still visible some places), but posterior part of metazona yellow, resulting in a banded look. Legs yellow. Preanal ring dark brown, anal valves yellowish brown, lighter towards the edges. Some individuals darker brown ranging to almost black (with greyish areas probably due to preservation). Yellow areas and colour pattern the same in all specimens. One specimen ( ZMUC 00020511) with a very light yellow colour instead of the brown colour; the yellow colour shared by all specimens being the darker of the two yellow nuances.
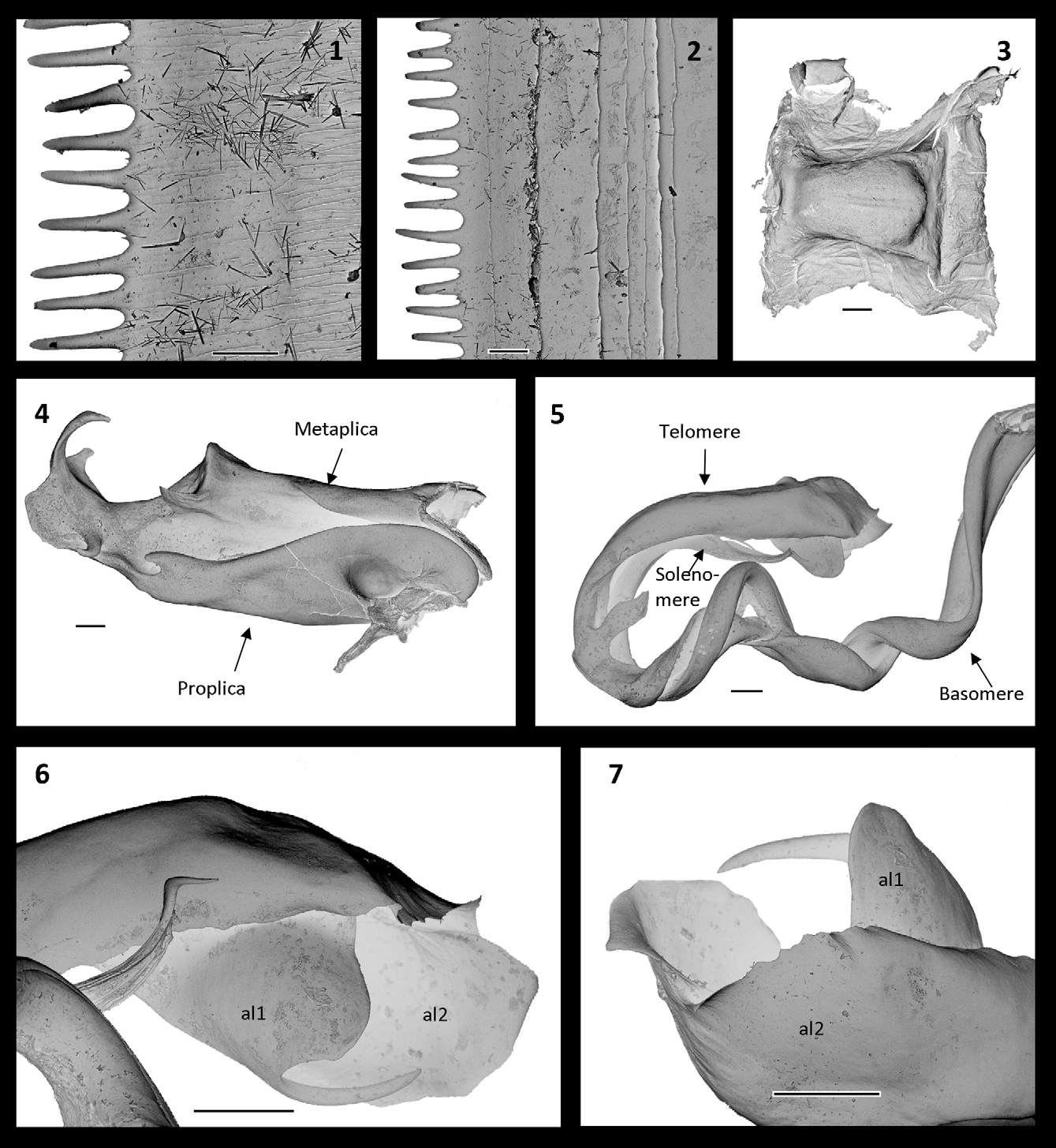
Head and body: 1+4+1 supralabral setae and a row of labral setae. Antennae reaching back to body ring 7. Ocelli in 6 horizontal rows. Length of eye (along upper row of ocelli) 0.53 × interocular distance. Distal margin of mandibular stipes shallowly bilobed. Lateral lobes of collum square, with three furrows, upper two furrows distinct, the lower marginal furrow barely visible. Ozopores from body ring 6, the exact location of the ozopores in relation to suture varying amongst individuals. Metazonites with furrows ventrally. Anal valves smooth, posteriorly perfectly rounded, each with a small dorsal spine. Anal valves apparently with at least one, possibly two posterior setae each. On many specimens the setae appear to be broken off. Limbus ( Fig.1–2 View FIGURE 1 – 7 ) serrated. Male legs with postfemoral and tibial pads.
Gonopods ( Figs. 3–10 View FIGURE 1 – 7 View FIGURE 8 View FIGURE 9 – 10 ): Anterior sternum as in X. robusta , a simple narrow triangle.—Coxa: Mesal metaplical flange (mmf) small as in all Xystopyge species. Proplica somewhat larger than mesal metaplical flange, but not as big as in other Xystopyge species, ending in a distad proplical lobe (pl). Metaplica with a broad flat rounded lateral metaplical process (lmp) with an anteriad lateral metaplical spine (lms) situated just below it, a pointed mediodistad apical metaplical process (amp), a triangular median metaplical process (mmp) projecting anteriad and forming a ‘roof’ over the emerging telopodite basomere, and a triangular distal metaplical flange (mf), situated between apical metaplical process and median metaplical process.—Telopodite: Basomere with an extended torsotope, without spines, distally slightly thickened. Solenomere whip-like with a spine apically (ass), closely appressed to telomere; solenomere and telomere forming a complete horizontal loop. Telomere ribbon-shaped, with a basal lamella (btl) ending distally in a blunt hook (h); c. 1/3 along its length a flattened, axehead-like process (tp) projecting into the telopodital loop. Telomere apically broadened, ending in two rounded lamellae (al1, al2. Fig. 6–7 View FIGURE 1 – 7 ); al1 with a spine-like process. Posterior sternum ( Fig. 3 View FIGURE 1 – 7 ) as in Xystopyge robusta : slightly elongate square, indented with a narrowing towards one end. There are minute variations in the exact shapes of the basal lamella , blunt hook and axehead-shaped telomeric process.
Remarks: The new species very much resembles the taxon described as X. robusta hoffmani by Vandenspiegel & Pierrard (2004). At the same place where X. pelecys has an axehead-shaped process, hoffmani has a hook-shaped process. The basal telomeric lamella seen in X. pelecys is lacking in hoffmani , which instead has a serrated lamella situated medially on the lower side (inside the loop) running the length of the telomere. The metaplica of X. pelecys is a bit simpler than that of X. hoffmani , having no triangular distolateral metaplical flange ( Vandenspiegel & Pierrard 2004: “projection dentiforme”) by the lateral metaplical process nor any rounded mesolateral metaplical flange ( Vandenspiegel & Pierrard 2004: “projection lamellaire”) by the proplical lobe. Instead, X. pelecys has a lateral metaplical spine situated between the proplical lobe and the lateral metaplical process.
Instead of regarding X. pelecys as another subspecies of X. robusta , due to its similarity especially with X. r. hoffmani , we treat it as a full species. At the same time, we formally elevate X. robusta hoffmani Vandenspiegel & Pierrard, 2004 , to a full species: X. hoffmani Vandenspiegel & Pierrard, 2004 , n.stat. We adopt this splitter approach because 1) we do not recommend use of the subspecies category in a group as poorly known as Odontopygidae , 2) recent studies have shown that even very subtle morphological differences between millipede populations may be linked to considerable genetic distances (e.g., Frederiksen et al. 2012, Pimvichai et al. 2011a,b).


| ZMUC |
Zoological Museum, University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |