
Atractotomus Fieber, 1858 ( Stonedahl,
|
publication ID |
https://doi.org/10.1206/0003-0082(2004)464<0001:ROPRWD>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/421687D2-BA18-2B77-FD46-0EC77954FC3A |
|
treatment provided by |
Carolina |
|
scientific name |
Atractotomus Fieber, 1858 ( Stonedahl, |
| status |
|
Atractotomus Fieber, 1858 ( Stonedahl, View in CoL View at ENA
1990), Europiella Reuter, 1909 ( Schuh et al., 1995), or Plagiognathus Fieber, 1858 ( Schuh, 2001) , is nonetheless confused for
1 Research Associate, Division of Invertebrate Zoology, American Museum of Natural History.
2 c/o Canadian National Collection of Insects, Agriculture and AgriFood Canada, 960 Carling Avenue, Ottawa, Ontario K1A 0C6 (schwartzm@agr.gc.ca).
3 106 Briza Court, Bellingham, Washington 98226.
Copyright © American Museum of Natural History 2004
ISSN 00030082
the same reason. Inadequate diagnoses founded on variable characters have resulted in an unstable taxonomy with changeable species affiliation and generic status. The species we treat as Phoenicocoris were previously placed in several generic level taxa: Asthenarius Kerzhner, 1962, Phoenicocoris Reuter , Psallus Fieber, 1858 , Salicarus Kerzhner, 1962 , and Sthenarus Fieber, 1858 .
Reuter (1875) described the subgenus Plagiognathus ( Phoenicocoris) to include the type species, Capsus modestus MeyerDür, 1843 , then upgraded Phoenicocoris to generic status ( Reuter, 1876), and when revising the European fauna ( Reuter, 1878), synonymized Phoenicocoris with Sthenarus . Reuter’s concept for Sthenarus focused on a group of eight species: dissimilis Reuter, 1878 ; maculipes Reuter, 1878; modestus MeyerDür, 1843 ; ochraceus Scott, 1872; ocularis Mulsant and Rey, 1852; pusillus Reuter, 1878; roseri HerrichSchaeffer, 1838 ; and, the type species, rotermundi Scholtz, 1847.
This divergent assemblage of species, plus an additional seven, was the subject of a detailed revision by Wagner (1958). The subsequent disposition of these species is explained in the following paragraphs. Wagner retained a single genus, Sthenarus , and segregated the taxa into two subgenera, providing new status for Phoenicocoris . Wagner’s diagnosis relied on the presence of scalelike or sericeous setae in the vestiture, the contour of the clypeus, the sharpness of the transverse basal carina of the vertex, and the extent of the thickening of antennal segment 2. All of these structures are prone to considerable variation and, while useful to distinguish individual species, are not diagnostic at the generic level ( Stonedahl, 1990).
In preparation for a key to the European USSR ( Kerzhner, 1964), Kerzhner (1962), was confronted with Wagner’s polyphylectic amalgamation of species and organized them into five groups founded primarily on the structure of the male genitalia and host plant utilization rather than characters of ‘‘secondary taxonomic importance’’. He limited the concept of Sthenarus to the type species and albipilis Wagner, 1958. Presumably both species feed on Populus and possess large vesicae which may indicate a relationship to Psallus . A species from Texas, pubescens Reuter, 1876, described in Phoenicocoris , was later placed in Sthenarus ( Atkinson, 1890) where it remained in all subsequent catalogs ( Van Duzee, 1917; Carvalho, 1958; Henry and Wheeler, 1988; Schuh, 1995). This enigmatic species is unknown to us and, along with S. australis Reuter, 1904 , is currently considered incertae sedis (viz. Schuh, 1995). Three other species, which awkwardly resided in Sthenarus , are now accommodated by new genera: cuneotinctus ( Van Duzee, 1915) in Aurantiocoris Schuh and Schwartz, 2003 , mcateei ( Knight, 1927) in Sthenaropsidea Henry and Schuh, 2002 , and myersi ( Woodward, 1950) in Xiphoides Eyles and Schuh, 2003 .
Kerzhner (1962) erected the genus Salicarus to accommodate the Salix feeding type species, roseri . He diagnosed the genus by the scalelike setae being limited to the thoracic pleuron; however, many, if not all of the other eight species currently placed in Salicarus (Schuh, 1995; Kerzhner, 1997), have scalelike setae on the dorsum. The large Sshaped vesica with fused apical processes and the large secondary gonopore situated on the distal half of the vesica are also diagnostic for the genus. In addition to willows ( Salicaceae ), the known hosts for Salicarus spp. are Halimodendron and Quercus (Fagaceae) , and Perovskia (Lamiaceae) .
The subgenus Psallus ( Asthenarius) was erected by Kerzhner (1962) to accommodate the type species, ocularis, and other Quercus feeding species distributed in the Mediterranean Region. This nominal subgenus was subsequently synonymized with P. ( Phylidea ) Reuter, 1899 ( Kerzhner, 1962), and currently contains 26 species ( Kerzhner and Josifov, 1999). The group is recognized by a stout, small, oval, and dark habitus, pale tibiae with black basal spine spots, and most importantly, a large ventrally keeled, genital segment of the males, with a massive Cshaped vesica terminating in a dentate apical ring ( Kerzhner, 1962).
Kerzhner (1962) raised Phoenicocoris to generic status and diagnosed modestus , the type species and two species transferred from Psallus : obscurellus Fallén, 1829 (subgenus Stenopsallus Wagner, 1960 ) and strobicola Knight, 1923 on genitalic and host plant characters. The vesica is strongly Sshaped, twisted, with two long, thin, subparallel apices of the vesical strap and with the secondary gonopore far removed from the apex of the vesica. All three taxa feed on Pinus spp. (Pinaceae) . Sthenarus dissimilis was not included in the newly conceived Phoenicocoris by Kerzhner (1962) because the vesica did not conform completely to the form seen in modestus (see discussion in the species treatment of dissimilis below).
Stonedahl (1990) transferred nine North American species (eight from Lepidopsallus Knight, 1923 and one from Atractotomus ) to Phoenicocoris . One of these species, P. knighti Kerzhner and Schuh, 1995 , was subsequently transferred to Pinomiris Stonedahl and Schwartz, 1996 (see discussion in Kerzhner and Schuh, 1995, and Stonedahl and Schwartz, 1996). Previous to these actions Phoenicocoris was represented in the Nearctic Region by two species, the introduced, dissimilis , and the endemic, strobicola . Stonedahl (1990) also suggested that several Palearctic species of Phoenicocoris apparently were not congeneric with modestus . At the time of that revision there were seven Old World species (including dissimilis and modestus ) recognized in the genus: carbonarius (Horváth) , flagellatus (Wagner) , kyushuensis (Linnavuori) , obscurellus , and vidali (Lindberg) . Phoenicocoris flagellatus was transferred by Vinokurov and Kanyukova (1995) to Salicarus and is now considered a junior synonym of fulvicornis (Jakovlev) ( Kerzhner, 1997) ; opacus (Reuter) was transferred from Psallus by Kerzhner (1997); and, qiliananus Zheng (in Zheng and Li, 1996) and ryukyuensis Yasunaga, 1999 , were described as new species. Currently 18 nominal species are recognized in Phoenicocoris and these are the subject of our revision. As a consequence of examining coniferinhabiting Phylini for several projects ( Stonedahl, 1990; Stonedahl and Schwartz, 1996; Schwartz and Schuh, 1999; Schwartz and Scudder, 2000, 2003) we have encountered specimens which require description of three new species of North American Phoenicocoris .
Specimen measurements (in millimeters) given are the mean and range (in parentheses) for 10 specimens of each sex, taken from across the distributional range, unless otherwise indicated. Scanning electron micrographs are of males. Micrographs of scalelike setae are taken from a dorsal view on the corium, unless otherwise noted. The localities of taxa used for comparison with Phoenicocoris are given in the figure captions or appendix. Descriptions and redescriptions are of males, with relevant female attributes not ed. Nomenclature for conifers conforms to the Gymnosperm Database ( Earle, 2002).
The generic descriptions include documentation of the female genitalia. Terminology for these structures roughly follows Davis (1955) and Slater (1950). We make reference to a particular modification of the base of the first valvulae and vestibulum which was first noted by Henry and Schuh (1979) and discussed by Eyles and Schuh (2003).
GENUS PHOENICOCORIS REUTER
Plagiognathus ( Phoenicocoris) Reuter, 1875: 99 (new subgenus). Type species: Capsus modestus MeyerDür, 1843 View in CoL (by monotypy).
Phoenicocoris: Reuter, 1876: 90 View in CoL (upgraded to genus); Kerzhner, 1962: 231 (new status, diagnosis, discussion); Schuh, 1995: 373 (catalog); Kerzhner and Josifov, 1999: 387 (catalog); Maw et al., 2000: 123 (checklist).
Stenopsallus Wagner, 1960: 12 (as subgenus of Psallus Fieber View in CoL ). Type species: Phytocoris obscurellus Fallén, 1829 View in CoL (by original designation); Kerzhner, 1962: 381 (synonym).
DIAGNOSIS: Recognized among other Phylini by the following combination of features: densely distributed white or silvery, scalelike setae on a variable portion of the dorsum (figs. 10A, 14A, 16A), and sometimes thoracic pleura (figs. 12A, 15A, 18A, 19A) and lateral margins of abdomen; labium long, almost always reaching the abdominal sternum; antennal segment 2 usually cylindrical in both sexes; distal onethird to onehalf of hind femur with irregular row of spinules dorsally (figs. 9C, 11B, 13B, 15D); pretarsus with slightly curved claw, moderately large pulvillus covering, at most, half of ventral surface of claw, and long apically spatulate parempodia (figs. 8C, 9C, 11A, 12C, 13C, 14D, 15C, 16D, 18C, 19E); and, vesica an Sshaped, strongly coiled strap, which is bifurcate distally, terminating in two long, usually divergent apices, with the secondary gonopore situated slightly distal to the medial coil, and with a variable length gonopore sclerite (figs. 11C, 19F, 24, 25).
REDESCRIPTION: Male: GENERAL AS PECT: macropterous, slightly to moderately elongate, total length 2.15–3.70; length from apex of clypeus to cuneal fracture 1.56–2.70; width across humeral angles of pronotum 0.98–2.15; yellowish brown, reddish brown, or brownish black general coloration; antennae, tarsi, tibiae, and apices of femora sometimes pale yellow to dusky brownish yellow; dorsum with dark brown to black simple setae and moderately to densely distributed, silvery white or white, lanceolate to ovate, adpressed, scalelike setae; thoracic pleura and lateral margins of abdomen sometimes with scalelike setae; pronotum sometimes without scalelike setae. HEAD: slightly to moderately produced anteriorly, posterior margin mostly straight; frons strongly sloping; vertex weakly convex, sometimes with low, broad transverse basal carina; maxillary plate slightly sunken; buccal cavity narrow, obovate; eyes occupying about onehalf to threequarters head height in lateral view, posterolateral margin contiguous with anterolateral margin of pronotum, broadly emarginate anteriorly; antennal insertion slightly dorsal to, or level with, ventral margin of eye; antennae pale yellow to dark brown, segment 1 short, length less than onehalf eye height; segment 2 uniformly cylindrical or sometimes slightly thickened distally, or rarely more strongly thickened and weakly fusiform; diameter of distal segments much less than segments 1 or 2; labium usually reaching beyond apices of hind coxae to base of genital segment. THORAX: pronotum trapezoidal, width about two times length, broadly rounded posterolaterally, lateral and posterior margins nearly straight; disk weakly convex, without distinct anterior and posterior lobes; calli weakly differentiated; mesoscutum moderately to broadly exposed; scutellum flat. HEMELYTRA: subparallel or weakly curved laterally; cuneus as long as broad; membrane moderately suffused with brown; veins dark. LEGS: coxae, trochanters, and most of femora usually dark, apices of femora, tibiae, and basal segment of tarsi pale; sometimes entire femora pale; femora without scalelike setae or dark spots; tibial spines black, usually without black basal spots. GENITALIA: genital segment relatively small, apex pointed or somewhat truncate; phallotheca relatively long apically, strongly curved; vesica with single sclerotized strap, coiled or strongly twisted medially, bifurcating at point even with distal end of secondary gonopore, with apices diverging distally, sometimes apex of anterior strap much shorter than apex of posterior strap; secondary gonopore situated much nearer medial coil than apex of strap, sometimes proximal portion of gonopore subtended by variable length gonopore sclerite; sometimes distal end of gonopore with spinules; right paramere simple; left paramere typically boatshaped.
Female: Macropterous, usually slightly more rounded than male, total length 2.30– 3.65; length from apex of clypeus to cuneal fracture 1.46–2.60; width across humeral angles of pronotum 0.98–2.16; similar to male except eyes slightly smaller, vertex wider, diameter of antennal segment 2 smaller, and labium reaching base of ovipositor. GENI TALIA: sclerotized rings subovoid, moderately large, folded, separated, mesial portion somewhat produced, ring widest at anterolateral margin; ventral labiate plate small, present on lateralmost portion of rings, produced dorsally adjacent to ring; dorsal structure large, membranous, spanning and broadly attached to interramal sclerites, concave anteriorly, sinuate dorsally, covered with field of minute spinules; vestibulum large, strongly sclerotized and convoluted, Sshaped, asymmetrical, strongly projecting anterior to right ring.
DISCUSSION: Our concept of Phoenicocoris is founded primarily on characters presented in the generic diagnosis. We will now present arguments for the inclusion of these defining features and discuss comparative information for related genera where possible.
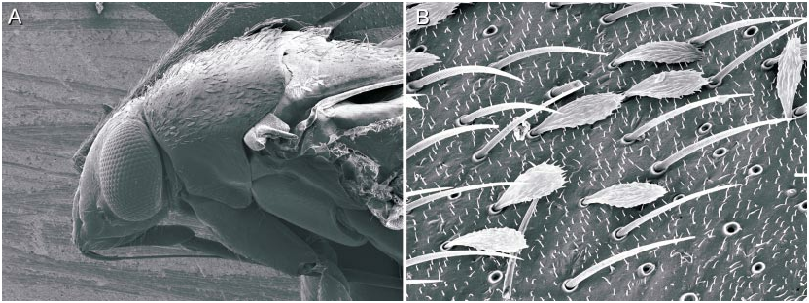
Sexual dimorphism in Phoenicocoris is limited to the type observed in the vast majority of plant bug species. The females have the mesial hemelytral width slightly greater and the lateral margins slightly more curved than in males. No species has shortened forewings in either sex. The eyes of the male are usually larger, in dorsal and lateral views, and accompanied by a narrower interocular width of the vertex; in females the eyes are smaller and the vertex is wider. Antennal segment 2 is of uniform diameter throughout its entire length in all Phoenicocoris spp. Only in minusculus is segment 2 of both sexes slightly fusiform. Segment 2 of Atractotomus mali is strongly fusiform and slightly flattened in both sexes. The length of segment 2 is roughly similar among the sexes in all the species treated herein.
The labium is generally long in Phoenicocoris with the apex reaching from the apices of the hind coxae to the middle of the abdominal venter or as far as the genital segment in males or the base of the ovipositor in females. Only in dissimilis , minusculus , and A. mali is the labium shorter, reaching only to the apices of the middle coxae.
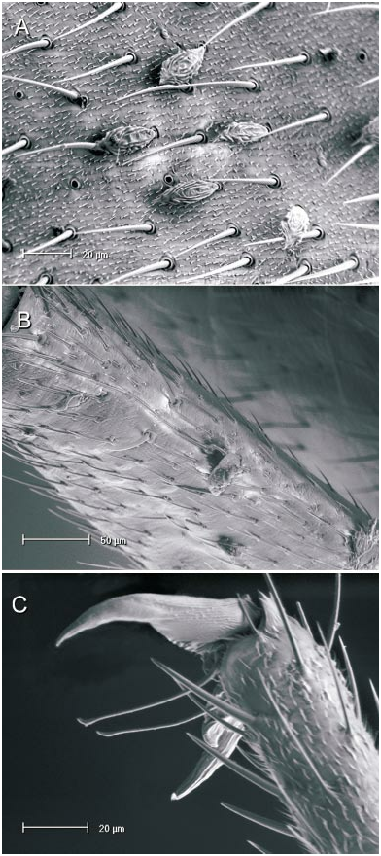
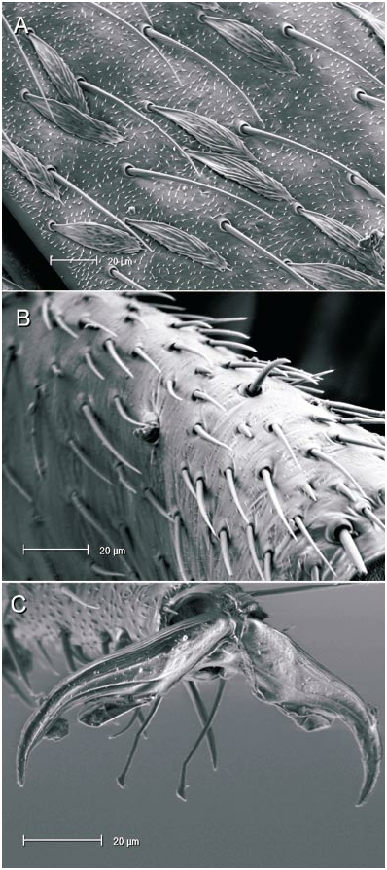
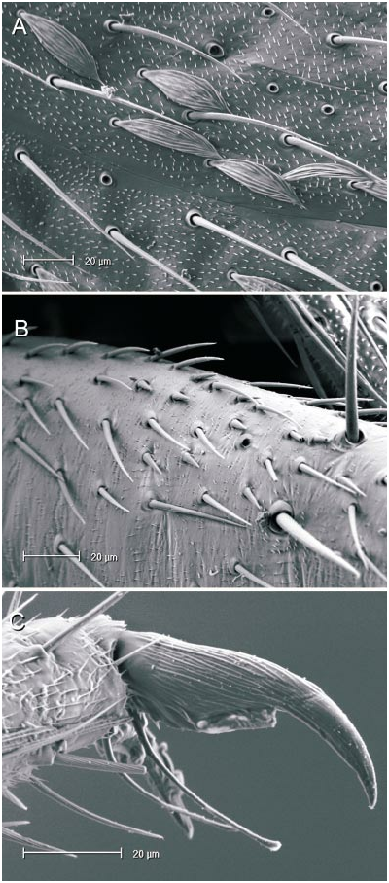
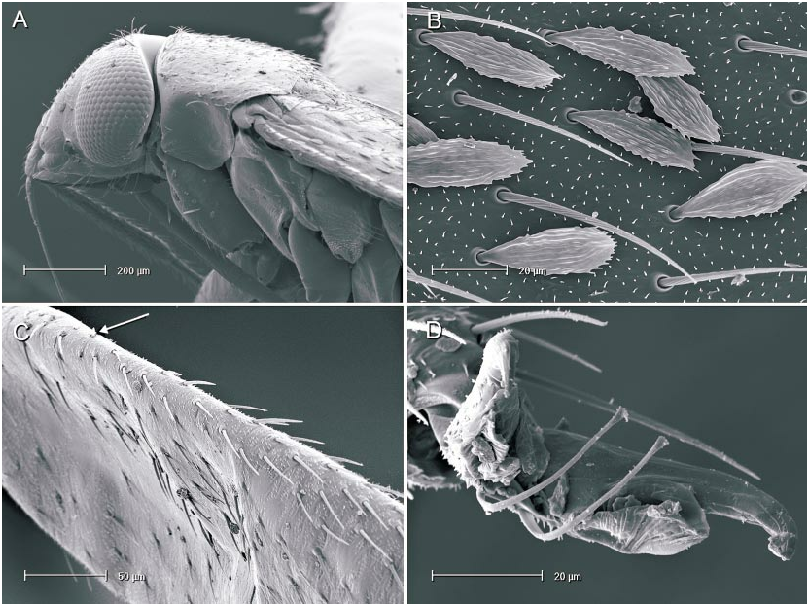
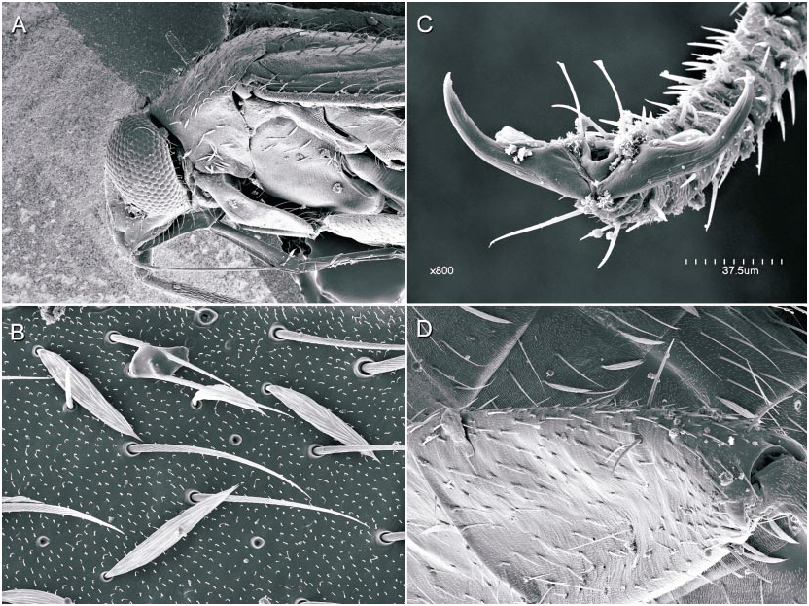
The microstructure of scalelike or sericeous flattened setae has been documented and described for a number of North American Phylini ( Schuh and Schwartz, 1985; Stonedahl, 1990; Stonedahl and Schwartz, 1996; Schuh, 2000a, 2000b, 2001). These authors have also defined several types of scalelike setae, discussed the utility of including vestiture features in defining monophyletic groups for the tribe, and incorporat ed setal characters in phylogenetic analyses. Schuh and Schwartz (1985) characterized two basic types of scalelike setae: (1) [narrow,] swollen mesially, somewhat flattened, and apically acuminate; and (2) [broad,] strongly flattened, apically broad, and ‘‘truly scalelike’’. Stonedahl (1990) referred to these two basic groups as types 1 and 2. Type 2 setae were further classified by these authors based on the ridge structure: (2a) distinctly raised, more or less parallel, which sometimes ramify apically; (2b) nearly parallel, but faint or totally obliterated, especially near the setal midline and apex; and (2c) midline of seta with noticeably anastomosed pattern. The scalelike setae of the taxa considered herein can likewise be placed in these categories but, as noted for other genera by previous workers, not all Phoenicocoris spp. have setal types identical to the type species.
The vestiture of almost all the Phoenicocoris spp. , as well as Atractotomus magnicornis (Fallén) , A. mali (MeyerDür) incertae sedis, and Salicarus roseri (HerrichSchaeffer) , are documented for the current study. Only the obscure Palearctic species opacus and vidali were not examined. Both types of scalelike setae were observed among examined taxa. As previously noted ( Schuh and Schwartz, 1985: fig. 51; Stonedahl, 1990: fig. 65) A. magnicornis has type 1 setae, but the more parallelsided, apically truncate setae shown in figure 4B, C demonstrate previously unreported variation. Salicarus roseri is the only species in which simple, unexpand ed setae were found on the dorsum (figs. 1B, 5D) and type 1 setae (figs. 1E, 5C) on the thoracic pleura. Type 1 setae were observed in dissimilis (figs. 1A, D, 20B), kyushuensis (figs 1G, 12B), obscurellus (figs. 1M, 15B), and strobicola (figs. 1N, 19B). The setae of all four species vary slightly in mesial width, but all have silvery coloration. The remain der of the Phoenicocoris species and A. mali have type 2 setae which vary in ridge structure, mesial width, and apical attenuation, but have similar white coloration. Type 2a setae with mostly complete ridges, moderate mesial width, and pointed apices are noted for claricornis (figs. 1O, 9B), modestus (figs. 1L, 13A), pallidicornis (figs. 1P, 16B), and ponderosae (figs. 1Q, 17A). Atractotomus mali has a more broad, apically truncate type 2a setae (figs. 1H, 7B) than the Phoenicocoris spp. Type 2c setae with anastomosed ridges, wide mesial width, and relatively broad, serrate apices were found in australis (figs. 1R, 8A), longirostris (figs. 1S, 10B), nevadensis (figs. 1T, 14B), and rostratus (figs. 1U, 18B; also see Stonedahl, 1990: fig. 59). As documented by Stonedahl (1990: fig. 58 for the junior synonym, crataegi ), only minusculus (figs. 1K, 12B) has type 2b setae with diminished ridges mesially, wide mesial width, and broadly truncate, serrate apices. For comparison we include documentation for the scalelike setae of three additional phyline species: Pruniocoris stonedahli Schuh and Schwartz with type 2a setae (fig. 1F); Arctostaphylocoris manzanitae Schuh and Schwartz with type 1 setae (fig. 1I); and, Guetherocoris atritibialis (Knight) with very narrow type 1 setae (fig. 1J).
Scalelike setae are not uniformly distributed across the body of the examined species, and this variation is incorporated in our key to the species of Phoenicocoris . In minusculus , obscurellus , rostratus , and strobicola , as well as Atractotomus magnicornis and A. mali , scalelike setae are found on the head, pronotum, thoracic pleura and abdominal venter. The reminder of the species do not have scalelike setae on the thoracic pleura, and abdominal venter, with modestus , nevadensis , and ponderosae also lacking scalelike setae on the pronotum. Although Wagner (1958) maintained that narrow scalelike setae are present on the dorsum of Salicarus roseri, Putshkov (1977) , Kerzhner (1962), and our observations (figs. 1B, 5D) indicate otherwise. Scalelike setae are present on the dorsum of other species of Salicarus (see fig. 2, for fulvicornis ), and all the species placed in Salicarus , including roseri , have scalelike setae on the thoracic pleura.
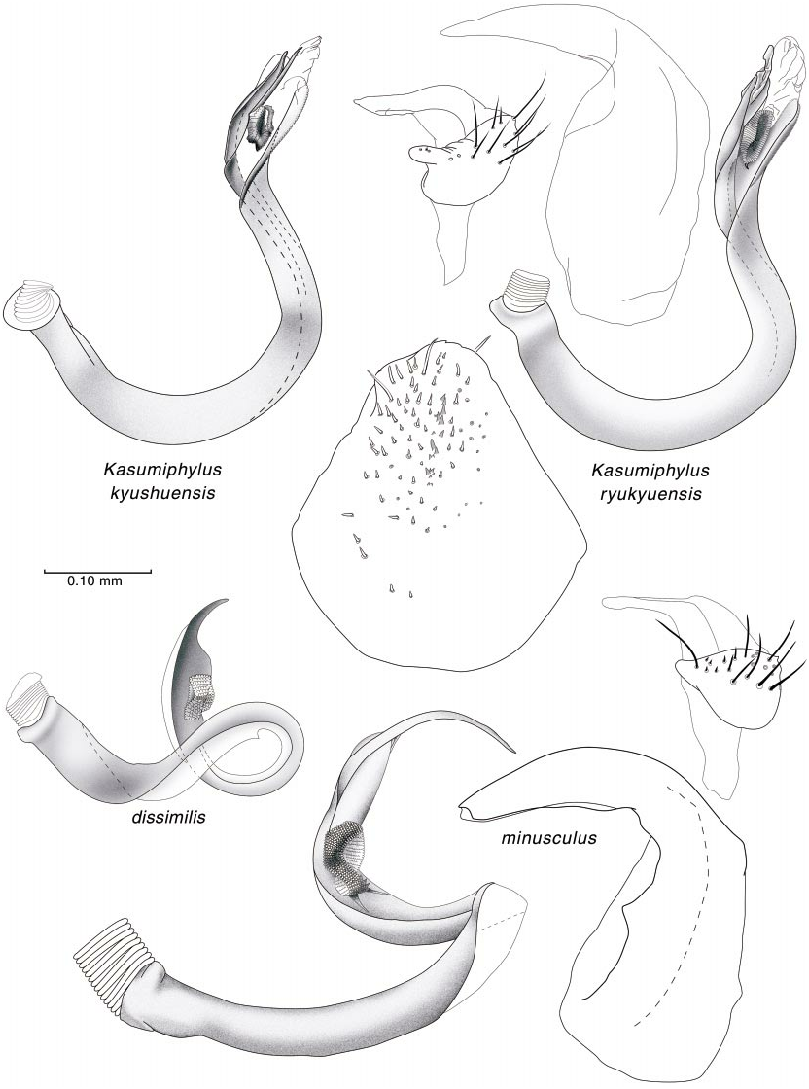
Hind femoral spicules were incorporated in the diagnoses of Rhinacloa Reuter 1876 ( Schuh and Schwartz, 1985), in which a nearly linear row of closeset spicules occurs, and Atractotomus ( Stonedahl, 1990) , in which a partial row of more widely spaced spicules is present. Stonedahl (1990: figs. 76, 78) also documented a shorter, less regular or dispersed row of spicules in two species of Phoenicocoris . In this present work, we confirm that all species of the genus have at least a few spicules placed somewhere on the hind femora (figs. 14C, 16C, 19D), if not a short group or row of distal spicules (figs. 8B, 9C, 11B, 12D, 13B, 15D, 16C, 17C, 18D). Furthermore we document that kyushuensis , which is now accommodated in the new genus Kasumiphylus (fig. 22D), and Salicarus roseri have several widely dispersed spicules on the hind femora (fig. 6B). Atractotomus mali apparently does not possess spicules on the distal portion of the hind femora (fig. 7C).
The value of including pretarsal microstructure in revisional studies of phyline plant bugs was initiated by Schuh (1976). Schuh and Schwartz (1985) and Stonedahl (1990) surveyed the pretarsal structure of many phyline, species providing a context in which we can interpret our observations. The claws of Phoenicocoris species are moderately curved in lateral view and have relatively broad bases and few, if any, claw hairs on the outer surface. The pulvillus is uniformly small throughout the genus, terminating mesial to the pronounced curve in the claw, and not extending ventrally more than a third of the claw height distal to the bend. The most striking feature of the Phoenicocoris pretarsus is the long parempodia, which terminate in a weakly spatulate apices. Parempodia with similar structure have been documented previously ( Schuh and Schwartz, 1985: fig. 73; Stonedahl, 1990: figs. 97, 99; Schuh, 2000b: fig. 20D) in several seemingly unrelated species: Campylomma verbasci (MeyerDür) , Heterocapillus genistae (Lindberg) , and Megalopsallus punctipes (Knight) . Now, A. mali and probably S. roseri are also known to possess apically spatulate parempodia (figs. 6A, 7D). A species of Lasiolabops Poppius 1914 , a member of the Leucophoropterini from the IndoPacific Region, demonstrates that long parempodia (however in this case, curved medially) with lamellate apices are not restricted to the Phylini ( Schuh, 1984: fig. 458). Phoenicocoris dissimilis does not have the same parempodial structure as its congeners. The parempodia of this species are relatively short, uniformly wide throughout, strongly lamellate, and apically truncate (figs. 21B, 21C). Such structure is unlike the majority of phylines, except Semium hirtum Reuter ( Schuh, 1976: fig. 27) and S. subglaber Knight (fig. 21D). However, lamellate parempodia exhibiting considerable structural diversity are well documented in the Pilophorini .
All species of Phoenicocoris have the femora concolorous with the dorsum, ranging from yellowish brown to reddish brown or brownish black; the tarsi, tibiae, and apices of the femora are pale yellow to dusky brownish yellow. Only in dissimilis , minusculus , and A. mali does the leg color differ from this pattern. In dissimilis the legs are dark brown to black with the apices of the femora and the basal tarsal segments dusky yellowish brown. The femora and tibiae are uniformly dark reddish brown to nearly black with the tarsi dusky yellow in minusculus . In A. mali the legs are dark reddish brown to black with the majority of the distal portion of the tibiae dusky yellowish or reddish brown.
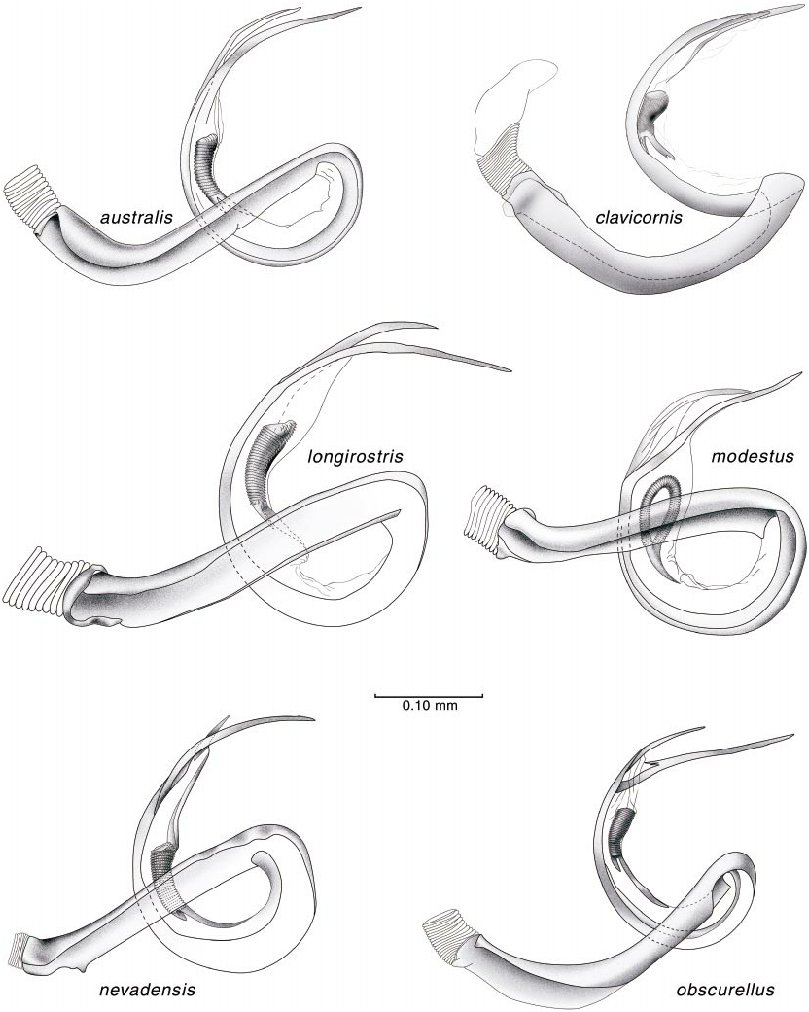
Male genitalia have been used extensively to define genera, species groups, and broader phylogenetic relationships in the Phylini . The generic concepts of Atractotomus ( Stonedahl, 1990) and Plagiognathus ( Schuh, 2001) were primarily constructed of male genitalic features, especially those of the vesica. We find that the vesical form, unlike the other characters discussed above, does not occur in taxa outside of what we diagnose as Phoenicocoris . Our investigation of the taxa historically placed in Phoenicocoris reveals vesical forms which corroborate some of the groups recognized by Kerzhner (1962) in his treatment of Sthenarus of European Russia. Species conforming to our conception of Phoenicocoris have a tightly coiled, single vesical strap which bifurcates at a point even with the base of the secondary gonopore. The apices of the strap usually diverge distally, with the anterior apex, which originates nearest the aperture of the secondary gonopore, the longer of the two apices; sometimes the anterior apex is shorter than the posterior apex (figs. 23, for nevadensis , 25, for minusculus ). The secondary gonopore is situated in the middle and ventral side of the vesical strap, just distal to the medial coil. A gonopore sclerite, originally known from a number of other phyline genera ( Atractotomus [ Stonedahl, 1990], Knightomiroides and Pinomiris [ Stonedahl and Schwartz, 1996], and Megalopsallus [ Schuh, 2000b]) is present in Phoenicocoris in which this variable length sclerite is devoid of surface spinules.
Species placed in Salicarus have a vesical structure quite similar to that of Phoenicocoris spp. The vesicae of S. fulvicornis and S. roseri , the species we examined in detail, are slightly larger and not as tightly coiled as in Phoenicocoris (fig. 26). The strap is singular and bifurcate, but the apices are fused along practically their entire length, and the secondary gonopore is placed closer to the apex of the strap. The vesica of dissimilis is tightly coiled, but its single strap is undivid ed distally and the apex is strongly sclerotized and conspicuously notched (fig. 25). The secondary gonopore does not have a subtending gonopore sclerite. Two Asian species, kyushuensis and ryukyuensis , previously placed in Phoenicocoris and now comprising Kasumiphylus , n.gen., have male genitalia which are of an entirely different form than that found in the three groups described above (fig. 25). The vesicae of these species are Jshaped, apically bifurcate, with roughly similar length apices, and have a terminal membrane. The secondary gonopore is situated subapically between the strap apices and the gonopore sclerite is absent. Uniquely in Kasumiphylus , the ventral surface of the genital segment is covered with stout bristles (fig. 25 for ryukyuensis ), a feature not seen in species of Phoenicocoris .
We treat 12 Phoenicocoris species in the following key, recognizing nine North America species, three of which, nevadensis , pallidicornis , and ponderosae , are new, and also include three Palearctic species, one of which, dissimilis , was introduced into the eastern United States.
BIOLOGY: Almost all the collection records and field observations indicate that conifers, predominately species of Pinus , are the breeding hosts for Phoenicocoris . Wheeler (2001) mentioned that some species of Phoenicocoris ‘‘develop on the new growth of conifers without inducing chlorosis or oth er obvious external symptoms on the needles’’. Only P. minusculus is consistently associated with nonconiferous hosts, being found on Crataegus and Malus , both genera of Rosaceae ; it is also known to be entomophagous ( Kelton, 1983). Plant hosts of the related taxa examined in this present study are either conifers or perennial shrubs of the same plant families discussed in Stonedahl (1990) for species of Atractotomus . Kasumiphylus spp. and P. dissimilis are conifer feeders (see species treatments below). Atractotomus mali prefers plant species from several rosaceous genera where it is both phytophagous and predaceous on aphids and mites ( Kelton, 1983). Wheeler and Henry (1992) listed apple ( Malus sylvestris Mill. ), crabapple ( Malus spp. ), hawthorn ( Crataegus oxycantha L.), pear ( Pyrus communis L.), plum ( Prunus spp. ), and scarlet firethorn ( Pyracantha coccinea M. J. Roem. ) as hosts of A. mali . Host records for the two species of Salicarus examined, fulvicornis and roseri , are from Caragana sp. (Fabaceae) ( Kerzhner, 1997) and white willow ( Salix alba L.) ( Salicaceae ) ( Wagner, 1975), respectively.
KEY TO SPECIES OF PHOENICOCORIS REUTER View in CoL
1. Thoracic pleura and lateral margins of abdominal sternum with woolly or scalelike setae (figs. 12A, 15A, 18A, 19A)..... 2
— Thoracic pleura and abdominal sternum without woolly or scalelike setae (figs. 10A, 14A, 16A).................. 5
2. Body sexually dimorphic—male elongate, female ovate (fig. 3); length of antennal segment 2 much greater than width of head across eyes; dorsum with narrow, lanceolate, somewhat woolly setae (figs. 1M, N, 15B, 19B)....................... 3
— Body not sexually dimorphic—both sexes ovate; length of antennal segment 2 less than to slightly greater than width of head across eyes; dorsum with broad, strongly adpressed, ovate scalelike setae (figs. 1O– U)............................. 4
3. Antennae and legs pale yellow; vesica as in figures 19F, 24; Nearctic Region........................ strobicola Knight View in CoL
— Antennae and legs yellowish brown to dark brown, tibiae sometimes slightly paler; vesica as in figure 23; Palearctic Region.................... obscurellus (Fallén) View in CoL
4. Antennal segment 2 in both sexes thickened, slightly fusiform (fig. 2), uniformly reddish brown to nearly black (fig. 2) scalelike setae on dorsum broadest apically, truncate (figs. 1K, 12B); vesica as in figure 25; eastern Nearctic Region.......................... minusculus (Knight) View in CoL
— Antennal segment 2 slightly thicker in male, cylindrical, yellow or yellowish brown (fig. 3); scalelike setae on dorsum broadest medially, pointed apically (figs. 1U, 18B); vesica as in figure 24; Canada, northern and western montane, United States...................... rostratus (Knight)
5. Pronotum without scalelike setae or with a few on anterior margin............. 6
— Pronotum with evenly distributed, flattened, scalelike setae.................... 8
6. Total body length greater than 3.0 mm; vesica as in figure 24; montane Arizona and Colorado........... ponderosae View in CoL , n.sp.
— Total body length less than 2.7 mm.... 7
7. Labium reaching well beyond apices of hind coxae, sometimes to eighth or nineth abdominal sternite in males, sometimes only to base of ovipositor in female; vesica as in figure 23; widely distributed in California, Nevada, and Oregon............................... nevadensis View in CoL , n.sp.
— Labium reaching to or just beyond apices of hind coxae; vesica as in figure 23; western Palearctic....... modestus (MeyerDür) View in CoL
8. Head, calli, anterior portion of pronotum, and most of mesoscutum and scutellum dark, posterior portion of pronotum and hemelytra noticeably paler; anteocular portion of head short (fig. 16A); vertex wide, approximately equal to onehalf width of head across eyes; vesica as in figure 24; central Canada..... pallidicornis View in CoL , n.sp.
— Head, pronotum, and hemelytra uniformly dull reddish brown, dark brown, or nearly black; anteocular portion of head prominent (figs. 10A, 20A); vertex more narrow, less than onehalf of width of head across eyes............................ 9
9. Antennal segment 1 reddish brown to nearly black; rarely segment 1 pale yellow (fig. 2, 3) but femora dark brown and from Sierra Nevada Mountains........... 10
— Antennal segment 1 pale to dusky yellow ............................... 11
10. Labium reaching posterior margin of mesosternum or sometimes middle of middle coxae, dorsum with narrow, lanceolate, woolly setae (figs. 1A, D, 20B); vesica as in figure 25; Palearctic Region and introduced in eastern United States........................ dissimilis (Reuter) View in CoL
— Labium reaching beyond apices of hind coxae, dorsum with broad, ovate, scalelike setae (figs. 1S, 10B); vesica as in figures 11C, 23; montane western United States............... longirostris (Knight) View in CoL
11. Femora uniformly pale to dusky yellow; vesica as in figure 23; Florida......................... australis (Blatchley) View in CoL
— Femora mostly darkened, only apices pale to dusky yellow; vesica as in figure 23; eastern United States................................ claricornis (Knight) View in CoL
Phoenicocoris australis (Blatchley) View in CoL
Figures 1R, 2 View Fig , 8 View Fig , 23 View Fig
Lepidopsallus australis Blatchley, 1926: 953 (n.sp.).
Phoenicocoris australis: Stonedahl, 1990: 60 View in CoL (n.comb.); Schuh, 1995: 374 (catalog).
DIAGNOSIS: Recognized by the uniformly chestnut or brownish red dorsum with broad, ovate, scalelike setae; pale yellow antennae and legs; thoracic pleura and lateral margins of abdominal sternum without scalelike setae; and by the secondary gonopore without prominent basal or apical spinules (fig. 23). Distribution of scalelike setae as in claricornis , dissimilis , longirostris , and pallidicornis . Distinguished from dissimilis and longirostris by the paler antennal segments 1 and 2 whereas these species have much darker antennal segments. The uniformly paler femora will distinguish australis from claricornis , which has only the apical portion of the femora pale, while the remainder of the femora is dark brown. Distinguished from pallidicornis by the uniformly reddish brown dorsum, whereas in pallidicornis the hemelytra, and usually a variable posterior portion of the pronotum, is obviously paler than the remainder of the dorsum.
REDESCRIPTION: ( N = 63, 8♀) GENERAL ASPECT: ovate; total length male 3.19 (3.00–3.40), female 3.19 (3.05–3.45); length to cuneal fracture male 2.26 (2.20–2.33), female 2.31 (2.21–2.43); coloration uniformly chestnut or reddish brown; antennae and legs, except for coxae, pale yellow and apex of tarsal segment 3, brown; dorsum, including vertex and pronotum, with moderately to densely distributed, broad, flattened, apically serrate, white scalelike setae and densely distributed, suberect to reclining, golden simple setae; thoracic pleura and venter without scalelike setae. HEAD: width male 0.74 (0.69–0.76), female 0.77 (0.74–0.79); vertex width male 0.31 (0.30–0.33), female 0.35 (0.32–0.36); region anterior to antennal insertion relatively short, anteocular length male 0.20 (0.19–0.21), female 0.22 (0.20– 0.23); eyes moderately large, about 83% of head height, ventral margin of antennal insertion dorsal to ventral margin of eye; antennal measurements male 0.19 (0.18–0.20): 0.82 (0.80–0.86): 0.44 (0.40–0.46): 0.32 (0.29–0.33), female 0.20 (0.19–0.20): 0.75 (0.72–0.78): 0.46 (0.44–0.50): 0.35 (0.34– 0.36); antennal segment 2 is 10% longer than head width across eyes; labium with basal segment brown, distal segments dusky yellow, reaching just beyond apices of hind coxae in males and reaching apex of sternite 6 in females; labial length male 1.56 (1.51– 1.60), female 1.69 (1.63–1.73); length of segment 4 male 0.45 (0.43–0.50), female 0.47 (0.44–0.49). THORAX: width male 1.09 (1.08–1.11), female 1.12 (1.08–1.14); length male 0.50 (0.49–0.53), female 0.51 (0.49–0.53); mesoscutum broadly exposed. HEMELYTRA: maximum width male 1.29 (1.25–1.34), female 1.38 (1.33–1.50); lateral margin slightly arcuate.
BIOLOGY: Collected on the microsporangiate strobili (‘‘male cones’’) of the presumed breeding host, slash pine ( Pinus elliottii Englemann ), where the male flower buds or bud stems did not appear to be injured (Edel, 1963).
DISTRIBUTION: From northern and west central Florida.
SPECIMENS EXAMINED: USA: Florida: Bradford Co.: R746 (10), [Hopk. U. S. 40181], 19.II.1960, B. H. Ebet, 3 flowers clusters, Pinus elliotti , reared 14.III.1960, 3♀ (USNM); R 770 (4), [Hopk. U. S. 40181], 14.II.1960, B. H. Ebet, 3 flower clusters, P. elliotti , reared 15.II.1960, 13, 5♀ (USNM). Columbia Co. : ‘‘Wiggen’s Place’’, Rd 450, [Hopk. U. S. 40181], 10.II.1960, B. H. Ebet, 3 flowers clusters, P. elliotti , reared, 23.II.1960, 43, 5♀ (USNM). Lake Co.: Lake City, 6.I.1958, B. H. Ebet, staminate flowers, P. elliotti , nymphs present, 23 (USNM).
Phoenicocoris claricornis (Knight)
Figures 10 View Fig , 2 View Fig , 9 View Fig , 23 View Fig
Lepidopsallus claricornis Knight, 1923: 471 (n.sp.).
Phoenicocoris claricornis: Stonedahl, 1990: 60 View in CoL (n.comb.); Schuh, 1995: 374 (catalog); Wheel er, 1999: 240 (biology).
DIAGNOSIS: Distinguished by the dark reddish brown to nearly black dorsum with broad, scalelike setae; antennae uniformly pallid to dusky yellowish brown; legs with femora mostly darkened, apices of femora, tibiae, and tarsi pale yellow; thoracic pleura and abdomen without scalelike setae; and by the contiguous apices of the vesical strap (fig. 23). Distribution of scalelike setae as in australis , dissimilis , longirostris , and pallidicornis . Distinguished from australis by the uniformly pale femora of this species, whereas in claricornis only the apical portion of the femora is pale, while the remainder of the femora is dark brown. Distinguished from dissimilis and longirostris by the paler antennal segments 1 and 2 whereas these species have much darker antennal segments. Additionally, in dissimilis the labium reaches only as far as the middle coxae; in claricornis the labium reaches the abdominal sternum. The uniformly dark dorsum of claricornis will distinguish this species from pallidicornis , which has a contrasting colored dorsum.
REDESCRIPTION: ( N = 93, 7♀) GENERAL ASPECT: small; total length male 2.81 (2.40–3.13), female 2.91 (2.73–3.00); length to cuneal fracture male 2.00 (1.85–2.13), female 2.13 (1.90–2.20); coloration dark reddish brown to nearly black; antennae, apices of femora, tibiae, and tarsi pale yellow to dusky brownish yellow; dorsum, including vertex and pronotum, with moderately to densely distributed, broad, flattened, apically pointed, white scalelike setae and densely distributed, suberect to reclining, somewhat unkempt, brown simple setae; thoracic pleura and venter without scalelike setae. HEAD: width male 0.72 (0.68–0.77), female 0.75 (0.72–0.78); vertex width male 0.31 (0.29– 0.33), female 0.34 (0.33–0.36); region anterior to antennal insertion relatively short, anteocular length male 0.19 (0.19–0.21), female 0.21 (0.20 –0.22); eyes moderately large, about 85% of head height, ventral margin of antennal insertion just dorsal to ventral margin of eye; antennal measurements male 0.18 (0.18–0.19): 0.75 (0.69 –0.81): 0.40 (0.34–0.43): 0.29 (0.26–0.31), female 0.17 (0.16–0.18): 0.70 (0.64–0.74): 0.41 (0.38– 0.48): 0.32 (0.28–0.38); segment 2 is 16% longer than head width across eyes; labium brown, reaching sternite 6 in males and subgenital plate in females; labial length male 1.44 (1.38–1.48), female 1.53 (1.43–1.59); length of segment 4 male 0.40 (0.38–0.41), female 0.40 (0.39–0.44). THORAX: width male 1.07 (1.05–1.10), female 1.07 (1.05– 1.13); length male 0.47 (0.41–0.49), female 0.49 (0.45–0.51); mesoscutum broadly exposed. HEMELYTRA: maximum width male 2.00 (1.85–2.15), female 2.13 (1.90– 2.20); subparallel laterally.
BIOLOGY: Wheeler (1999) discussed the seasonal history, habits, and immature stages from throughout the range of this species. The host plants are pitch pine ( Pinus rigida Miller ) and Virginia or scrub pine ( P. virginiana Miller ) ( Pinaceae ) where immature and adult stages feed on microsporangiate strobili.
DISTRIBUTION: Originally described from New Jersey ( Knight, 1923) , then recorded from West Virginia ( Wheeler et al., 1983). Wheeler (1999) provided records which span the eastern United States east of, and within, the Appalachian Mountains from Maine south to Georgia and as far west as Nelson Co., Kentucky and Dekalb Co., Alabama .
SPECIMENS EXAMINED: USA: District of Columbia : 20.V.1905 and 8.VI.1905, 2♀ ( USNM). New York: Saratoga Co.: Co. airport, N of Ballston Spa , 12.VI .1993 , A. G. Wheeler, Jr., Pinus rigida , 13, 2♀ (USNM). Warren Co.: Queensbury , 12.VI .1993 , A. G. Wheeler, Jr., P. rigida , 13 (USNM). North Carolina: Wake Co.: Raleigh , 3.V .1969 , D. A. Young, 53 (NCSU). Pennsylvania: Dauphin Co.: Fishing Creek Valley School, Rte 443, 22.V.1976 and 30.IV.1976, A. G. Wheeler, Jr., 5th instar, P. virginiana , 13, 1♀ ( USNM). Tennessee: Cocke Co.: Rtes 25 and 70, betw. 107N and 107S , 20.V .1979 , Wygodzinsky, Schuh, Schmidt, 1♀ (AMNH). Virginia: Fairfax Co.: Falls Church, 2. X, N. Banks, 1♀ (USNM). Roanoke Co. : NE of Roanoke on I81, 16.V .1988 , M. D. Schwartz, P. virginiana , 13, 1♀ (AMNH).
Phoenicocoris longirostris (Knight)
Figures 1S, 2 View Fig , 10 View Fig , 23 View Fig
Lepidopsallus longirostris Knight, 1968: 54 (n.sp.).
Phoenicocoris longirostris: Stonedahl, 1990: 61 View in CoL (n.comb.); Schuh, 1995: 374 (catalog).
DIAGNOSIS: Distinguished by the broad, scalelike setae on the dorsum; thoracic pleura and abdominal sternum without scalelike setae; long labium, especially segment 4; long anteocular portion of head; antennal segments 1 and 2 usually brownish yellow to brown, length greater than width of head across eyes; legs with femora reddish brown to dark fuscous, tibiae dusky yellow or yellowish brown; and the secondary gonopore with distal spinules (fig. 23). Frequently collected with rostratus in montane western North America, but the absence of scalelike setae on the thoracic pleura readily distinguishes longirostris . Distinguished from nevadensis and ponderosae by the absence of scalelike setae on the pronotum whereas in longirostris the pronotum has evenly distributed scalelike setae.
REDESCRIPTION: ( N = 143, 11♀) GEN ERAL ASPECT: moderately elongate; total length male 3.32 (3.05–3.60), female 3.36 (3.05–3.50); length to cuneal fracture male 2.35 (2.25–2.60), female 2.46 (2.34–2.55); coloration dark brown to nearly black; apices of femora, tibiae and tarsi pale yellow to dusky brownish yellow; sometimes antennal segments 1 and 2 pale yellow (fig. 2, 3), bases of femora sometimes darker brown; dorsum with moderately to densely distributed, broad, flattened, apically serrate, white scalelike setae and densely distributed, suberect to reclining, black simple setae; thoracic pleura and venter without scalelike setae. HEAD: width male 0.73 (0.69–0.75), female 0.75 (0.73–0.76); vertex width male 0.33 (0.30–0.35), female 0.35 (0.34–0.36); region anterior to antennal insertion elongate, anteocular length male 0.25 (0.24–0.28), female 0.27 (0.26–0.28); eyes moderately small, 77% of head height, ventral margin of antennal insertion level with ventral margin of eye; antennal measurements male 0.20 (0.18–0.21): 0.76 (0.70–0.88): 0.47 (0.44– 0.55): 0.34 (0.33–0.35), female 0.21 (0.19– 0.25): 0.82 (0.79–0.91): 0.50 (0.45–0.53): 0.36 (0.33–0.39); antennal segment 2 equal to or as much as 5% greater than width of head across eyes; labium brown, reaching base of genital segment or ovipositor; labial length male 1.74 (1.64–1.84), female 1.79 (1.73–1.88); length of segment 4 male 0.52 (0.50–0.58), female 0.53 (0.51–0.54). THO RAX: width male 1.07 (1.00–1.15), female 1.11 (1.05–1.16); length male 0.52 (0.50– 0.55), female 0.51 (0.50–0.53); mesoscutum moderately exposed. HEMELYTRA: maximum width male 1.35 (1.30–1.43), female 1.50 (1.37–1.60); subparallel laterally.
BIOLOGY: Breeds on whitebark pine ( Pinus albicaulis Engelmann ), bristlecone pine ( P. aristata Engelmann ), lodgepole pine ( P. contorta Douglas ex Loudon and subsp. murryana (Balfour) Engelmann ), limber pine ( P. flexilis E. James ), and subalpine fir ( Abies lasiocarpa (Hooker) Nuttall ).
DISTRIBUTION: Southern Yukon Territory, British Columbia , western Alberta and Cypress Hills in Alberta and Saskatchewan; widely distributed in montane regions of western United States south to Arizona.
DISCUSSION: The color of antennal segments 1 and 2 in longirostris is variable. Specimens from the Sierra Nevada Mountains of California and Nevada and northern California have segments 1 and 2 which are apparently always pale yellow. Specimens from the remainder of the distributional range of this species have dusky brown to dark brown segments. Both longirostris and rostratus were collected together from Pinus contorta at the same locality near Washoe, Nevada. In addition to the distribution of scalelike setae on both species (only the thoracic pleura of rostratus with scalelike setae), the coloration of segment 1 is also diagnostic; in longirostris segment 1 is pale yellow and in rostratus segment is dark brown.
SPECIMENS EXAMINED: CANADA: Alberta: Banff–Jasper Highway, Banff National Park, 25.VIII.1979, L. A. Kelton, 1♀ (CNC); Coal Valley, 31.VIII, 1978, L. A. Kelton, P. contorta , 23, 1♀ (CNC); Crowsnest, 19.VII.1975, L. A. Kelton, P. contorta , 63, 10♀ (CNC). Cypress Hills Provincial Park: Elkwater Lake, 20.VII.1956, O. Peck, P. contorta , 1♀ (CNC); Elkwater Park, 21, 28.VII, 14, 16.VIII.1952, L. A. Konotopetz, P. contorta murryana , 123, 21♀ (CNC). Eisenhow er Junction, 5.VIII.1970, L. A. Kelton, P. contorta , 1♀ (CNC); Fisher Creek Forest Station, 26.VII.1973, L. A. Kelton, 23, 1♀ (CNC). Kananaskis Highwood/Cataract Zone, 24.VII.1994, M. D. Schwartz: Highwood House, just S of jct Hwy 40 and Rte 940, P. contorta , 13 (CNC); Lineham Creek Picnic Area on Hwy 40, ex Abies lasiocarpa , 1♀ (CNC). Kananaskis Road, 20, 25, 27.VII.1974, L. A. Kelton, P. contorta , 33, 5♀ (CNC). Lake Louise, 25.VII.1973, L. A. Kelton, P. contorta , 73, 5♀ (CNC); Waterton National Park, 4–6.VII.1970, 26.VII.1970, L. A. Kelton, P. contorta , 73, 8♀ (CNC). British Columbia : Aspen Grove, 25.VII.1970, L. A. Kelton, P. contorta , 13, 2♀ (CNC); Atlin, 15.VIII.1955, H. J. Huckel, 1♀ (CNC); Christina Lake, 12.VII.1970, L. A. Kelton, P. contorta , 13 (CNC); Fernie, 7.VII.1970, L. A. Kelton, P. contorta , 53, 9♀ (CNC); Fruitvale, 11.VII.1970, L. A. Kelton 1♀ (CNC); Jaffray, 8.VII.1970, L. A. Kelton, P. contorta , 13 (CNC); Kaslo, 10.VII.1970, L. A. Kelton, P. contorta , 2♀ (CNC); Moyie, 9.VII.1970, L. A. Kelton, (CNC). Manning Provincial Park, Blackwall, L. A. Kelton, Abies lasiocarpa : 8.VII.1970, 13 (CNC); 23.VII.1970, 23, 8♀; Blackwall Peak Area, parking lot, alpine meadows nr Naturalist Hut, 19.VIII.1998, M. D. Schwartz, A. lasiocarpa, (UBC) . Moyie, 9.VII.1970, L. A. Kelton, 1♀ (CNC); Nickel Plate City, Hedley, 5000 ft, 12.IX.1953, J. E. H. Martin, 1♀ (CNC); Summit, 16.VIII.1982, L. A. Kelton, P. contorta , 503, 41♀ (CNC); Terrace, W. R. Richards: 7.VII.1960, 13 (CNC); 17.VII.1960, 13 (CNC). Trail, 21.VII.1959, L. A. Kelton, 13 (CNC); Yahk, 22.VII.1959, L. A. Kelton, P. contorta , 13, 6♀ (CNC); Yoho National Park, 16.VIII.1970, L. A. Kelton, 13, 1♀ (CNC). Saskatchewan: Cypress Hills Provincial Park, 4.VII.1970, L. A. Kelton, P. contorta , 163, 30♀ (CNC). Yukon Territory: Carcross: 9.VIII.1983 and 1.VIII.1982, L. A. Kelton, P. contorta , Lupinus sp. , 223, 43♀ (CNC); 28.VI.1982, G. G. E. Scudder, P. contorta , listed in Scudder 1997, 23, 3♀ (UBC). Morley Lake, 9.VIII.1982, L. A. Kelton, P. contorta , 53, 8♀ (CNC); Moose Creek, 28.VII.1983, L. A. Kelton, Pinus sp. , 1♀ (CNC); Rancheria, 11.VII.1982, L. A. Kelton, 183, 28♀ (CNC); Squanga Lake, 7.VIII.1982, L. A. Kelton, P. contorta , 13, 2♀ (CNC); Watson Lake, 13.VIII.1982, L. A. Kelton, 13, 2♀ (CNC). Whitehorse, L. A. Kelton: 16.VII.1982, Picea sp. , 4♀ (CNC); 30.VII.1982, Pinus contorta , 23, 5♀ (CNC). USA: Arizona: Cochise Co. : Rustler Park, Chiricahua Mountains, 7, 8.VII.1968, L. A. Kelton, P. ponderosa , 43, 2♀ (CNC). California: Alpine Co.: Alpine Lake, W end, Toiyabe National Forest, on Rte 4, 8000 ft, 5.VII.1994, M. D. Schwartz, P. contorta , 13 (CNC). El Dorado Co.: Lake Tahoe, 8.VIII.1937, Drake, Andre, Lepidopsallus hesperus paratype, 1♀ (USNM); near Bijou, Lake Tahoe, 19.VII.1929, R. L. Usinger, 13, 1♀ (UCB). Mariposa Co. : Yosemite Creek Ranger Station, Yosemite National Park, 21.VII.1946, R. L. Usinger, P. murrayana , 1♀ (UCB). Lassen Co.: Bogard Campground, 24 mi W jct. Hwys. 36 and 44, 10.VIII.1980, G. M. Stonedahl, P. contorta , 23, 1♀ (AMNH); Martins Springs, T31N, R9E, 5.VIII.1922, J. O. Martin, 1♀ (CAS). Plumas Co.: Caribou, 6.VII.1932, R. L. Usinger, 23 (UCB). Siskiyou Co.: Medicine Lake, on Medicine Lake Rd, G. M. Stonedahl, J. D. McIver, P. contorta : 1.4 mi S of, 19.VII.1985, 13 (AMNH); 2.5 mi N of, 18.VII.1985 33, 5♀ (AMNH). 6.6 mi S of Lava Beds National Monument, Lava Beds–Medicine Lake Road, 18.VII.1985, G. M. Stonedahl, J. D. McIver, P. contorta , 33, 8♀ (AMNH). Tuolumne Co.: Yosemite National Park, McCabe Lakes, 2.VIII.1939, R. L. Usinger, 1♀ (UCB). Colorado: Boulder Co.: Nederland, Roosevelt National Forest, 4.VIII.1968, L. A. Kelton, P. albicaulis , 2♀ (CNC); Rainbow Lakes, Roosevelt National Forest, 9800 ft, 3.VIII.1968, L. A. Kelton, 1♀ (CNC). Clear Creek Co.: Clear Creek Picnic Ground, 9400 ft, 10.VIII.1987, D. A. Polhemus, P. flexilis , 13, 2♀ (JTP); Green Lake, Guanella Pass, 9900 ft, 6.IX.1982, D. A., J. T. Polhemus, P. aristata , 1♀ (JTP). Juniper Pass on Rte 103, 10,500 ft, 21.VIII.1986, R. T. Schuh, J. T. Polhemus, P. contorta , 13, 2♀ (AMNH). Mt. Goliath area, 11,200 ft, 21.VIII.1986, R. T. Schuh, J. T. Polhemus, P. aristata , 13 (AMNH). Squaw Pass, Arapaho National Forest, 9800 ft, 27.VII.1968, L. A. Kelton, P. contorta , 63, 14♀ (CNC); Squaw Pass Road, J. T., D. A. Polhemus: 8.VIII.1983, P. aristata , 13, 2♀ (JTP); 14.VIII.1982, P. contorta , 23 4♀ (JTP). West Chicago Creek Campground, Arapaho National Forest, 28, 9800 ft, 29.VII.1968, L. A. Kelton, P. contorta , 73, 11♀ (CNC). Gilpin Co.: Pinecliffe, 12.VIII.1973, J. C. Schaffner, 43, 2♀ (TAM); Rollinsville, Roosevelt National Forest, 1.VIII.1968, L. A. Kelton, P. contorta , 3♀ (CNC). Grand Co.: Grand Lake, 12.3 mi N of, on Hwy. 34, 16.VIII.1980, G. Stonedahl, P. contorta , 1♀ (AMNH); Grand Lake entrance, Rocky Mountain National Park, 18.VIII.1968, L. A. Kelton, P. contorta , 4♀ (CNC); St. Louis Creek Campground, 3.4 mi W of Fraser, 9000 ft, 16.VIII.1980, Stonedahl, P. contorta , 13, 4♀ (AMNH). Jackson Co.: Crowley, nr, at Jackson Co. and Wyoming State line on Rte 125, 4.VIII.1979, J. T. Polhemus, 13 (JTP); Gould, 5.VIII.1975, J. C. Schaffner: 2 mi E of, 63, 2♀ (TAM); 4 mi N of, 23, 1♀ (TAM). Gould, 9200 ft, 12, 13.VIII.1968, L. A. Kelton, P. contorta , 53, 11♀ (CNC). Larimer Co.: Chambers Lake, Roosevelt National Forest, 9200 ft, 11.VIII.1968, L. A. Kelton, P. contorta , 1♀ (CNC); Fish Creek Picnic ground, Pingree Park Road, 46 mi W of Fort Collins. 7700 ft, 14.VII.1986, R. T. Schuh, J. T. Polhemus, 23 (AMNH); Milner Pass, Rocky Mountain National Park, 10,500 ft, 18.VIII.1968, L. A. Kelton, 1♀ (CNC). Pingree Park: 20.VIII.1925, H. H. Knight, Lepidopsallus longirostris paratypes, 33, 11♀ (CNC, USNM); 25.VIII.1976, Beamer, Lawson, 13, 1♀ (USNM). Park Co.: Geneva Park, 9600 ft, 6.XI.1982, D. A., J. T. Polhemus, P. flexilis , 4♀ (AMNH, JTP). Routt Co.: Meadows, Routt National Forest, 20, 21.VIII.1968, L. A. Kelton, Picea sp. , 13 (CNC); Steamboat Springs, H. H. Knight: 7000 ft, 16.VII.1964, 2♀ (USNM); 8500 ft, 17.VII.1964, 13 (USNM). Summit Co.: 8.7 mi W of jct. Hwy 91 on I70, 14.VIII.1980, G. M. Stonedahl, Pinus contorta , 103, 6♀ (AMNH); Loveland Pass, 12,000 ft, 7.VIII.1961, C. M. Mann, 13 (CNC); Vail Pass summit, 10,350 ft, 11.VIII.1987, J. T., D. A. Polhemus, P. flexilis , 13 (JTP). Idaho: Benewah Co.: Charlie Creek Rd., 4 mi E of Emida, 8.VII.1979, G. M. Stonedahl, 23, 1♀ (AMNH); 4 mi W of Emida on Hwy 6, 9.VII.1979, G. M. Stonedahl, P. contorta , Pseudotsuga menziesii , 2♀ (AMNH). Bonner Co.: Pend Orielle River, Laclede, 4.VII.1966, W. Gagne, J. Haddock, Pinus flexilis , 3♀ (UCB). Franklin Co.: Strawberry Canyon, 0.5 mi N of m.p. 18 on ID Rte 36, T13S R14E Sec. 1, 8000 ft, 19.VII.1981, M. D. Schwartz, P. contorta , 43, 9♀ (AMNH). Idaho Co.: Lochsa River Valley, E of Wilderness Access Campground (m.p. 127), 2400 ft, 1.VIII.1987, G. M. Stonedahl, Picea sitchensis , 1♀ (AMNH); Lowell, Clearwater National Forest, 30.VII.1972, L. A. Kelton, 2♀ (CNC). Latah Co.: Palouse River, 5 mi E of Harvard, 3.VII.1966, W. Gagne, J. Haddock, 1♀ (UCB). Lemhi Co.: Meadow Lake Campground, 3 mi W of Gilmore, W of Rte 28, Lemhi Range, Salmon National Forest, 9600 ft, 31.VI.1994, M. D. Schwartz, Pinus albicaulis , 13 (CNC). Teton Co.: Pine Creek Pass, 6.VIII.1972, L. A. Kelton, P. contorta , 33, 3♀ (CNC). Montana: Deer Lodge Co.: Georgetown Lake on Rte 10A, 6500 ft, 9.VIII.1986, Schuh, Schwartz, Stonedahl, P. contorta , 1♀ (AMNH). Glacier Co.: Glacier National Park, 28.VII.1972, L. A. Kelton, P. contorta , 23, 5♀ (CNC). Park Co.: Colter Campground, 2 mi E of Cooke City on Rte 212, 8000 ft, 11.VIII.1986, Schuh, Schwartz, Stonedahl, P. contorta , 3♀ (AMNH); Soda Butte Campground, 2 mi E of Cooke City, 7700 ft, 11.VIII.1986, Schuh, Stonedahl, Schwartz, P. contorta , 1♀ (AMNH); Rte 212 at WY Border, 7750 ft, 11.VIII.1986, Schuh, Schwartz, Stonedahl, P. contorta , 1♀ (AMNH). MontanaIdaho: Rte 191, Targhee Pass, 7072 ft, 10.VIII.1986, Schuh, Schwartz, Stonedahl, P. contorta , 13, 3♀ (AMNH). Nevada: Washoe Co.: Little Valley Research Area, 4.5 mi SW of Washoe, T16N R19E Sec2 NE 1/4, 6200 ft, 4.VIII.1982, M. D. Schwartz, P. contorta , 43, 14♀ (AMNH). Oregon: Baker Co.: West Eagle Meadow, 20 mi E of Medical Springs, Wallowa Mts., 4700 ft, 3.VIII.1986, Schuh, Schwartz, Stonedahl, P. contorta , 43, 11♀ (AMNH). Clackamas Co.: 1 mi S of Government Camp, 25–29.VII.1966, W. Gagne, J. Haddock, P. contorta , 13, 1♀ (UCB). Grant Co. : Wildcat Springs Campground, Malheur National Forest, T14S, R33E, Sec. 10, 21.VII.1979, M. D. Schwartz, P. contorta , 33, 4♀ (AMNH). Hood River Co.: Mount Hood, Cloud Cap, 6000 ft, 5.IX.1979, G. M. Stonedahl, P. contorta , 1♀ (AMNH). Klamath Co.: 19 mi SE of LaPine on Hwy. 31, 4750 ft, 25.VI.1979, G. M. Stonedahl, P. contorta , 2♀ (AMNH); T30S R8E Section 9, 13.VII.1978, Oman, 23, 3♀ (OSU); Skookum Meadows, R9F, T27S, Section 16, 5310 ft, 17.VII.1979, G. M. Stonedahl, P. contorta , 1♀ (AMNH); Meadow Spring, Winema National Forest, T27S R9E S16, 5500 ft, 17.VII.1979, M. D. Schwartz, P. contorta , 13 (AMNH). Lane Co.: Frog Camp Campground, NE Corner, 2.VIII.1966, W. Gagne, J. Haddock, 2♀ (UCB). Umatilla Co.: Blue Mountains, 4.VII.1940, R. L. Usinger, 13 (UCB). Union Co.: 0.1 mi NE of Tollgate Shopping Center on Hwy 204, 17.VIII.1979, M. D. Schwartz, P. contorta , 1♀ (AMNH). Wallowa Co.: Miram Lk. Trail, Wallowa–Whitman National Forest, 5590–7370 ft, 18.VIII.1979, M. D. Schwartz, P. contorta , 1♀ (AMNH). Utah: Cache Co.: 14 mi S of FS Rd 055 off Rte 89, T13N R4E S15, 8000 –9000 ft, 25.VII.1981, M. D. Schwartz, P. contorta , 53, 1♀ (AMNH). Uintah Co.: nr Little Brush Creek, m.p. 22 on Rte 44, T1N, R22E, Uinta Mountains, 8620 ft, 2.VIII.1981, M. D. Schwartz, P. contorta , 1♀ (AMNH). Wyoming: Albany Co.: Medicine Bow Mountains, 29.VII.1931, H. H. Knight, Lepidopsallus longirostris paratype, 13 (USNM). Fremont Co.: Blue Ridge Summit, Wind River Mountains, Shoshone National Forest, 14 mi SW of National Forest Boundary on Rte 131, 9576 ft, 14.VIII.1986, Schwartz, Stonedahl, P. contorta , 13, 6♀ (AMNH). Park Co.: Yellowstone National Park, 4.VIII.1931, H. H. Knight, 1♀ (USNM). Shoshone Co.: Fox Creek Campground, 6.9 mi E of Cooke City on Rte 212, 7250 ft, 11.VIII.1986, Schuh, Schwartz, Stonedahl, P. contorta , 1♀ (AMNH). Teton Co.: Colter Bay Village, 2 mi SE of, on Rte 89, Grand Teton National Park, 7000 ft, 22.VII.1981, M. D. Schwartz, P. contorta , 13, 5♀ (AMNH); Fishing Bridge Residential Area, Yellowstone National Park, 8400 ft, 21.VII.1981, M. D. Schwartz, P. contorta , 13, 1♀ (AMNH); Grand Teton National Park, 7.VIII.1972, L. A. Kelton, P. contorta , 3♀ (CNC).
Phoenicocoris minusculus (Knight)
Figures 1K, 2 View Fig , 12 View Fig , 25 View Fig
Lepidopsallus minusculus Knight, 1923: 472 (n.sp.).
Phoenicocoris minusculus: Stonedahl, 1990: 61 View in CoL (n.comb.); Schuh, 1995: 374 (catalog).
Atractotomus crataegi Knight, 1931: 37 (n.sp.). NEW SYNONYM.
Phoenicocoris crataegi: Stonedahl, 1990: 61 View in CoL (n.comb.); Schuh, 1995: 374 (catalog).
DIAGNOSIS: Recognized by the uniformly dark reddish brown to nearly black dorsum with broad, truncate, apically serrate, white scalelike setae (fig. 12B) on the dorsum, thoracic pleura, and lateral margins of abdominal sternum (fig. 12A); the thickened, slightly fusiform antennal segment 2, in both sexes (fig. 2), with length less than to slightly greater than width of head across eyes (ratio male 1:1.04–1.13; female 1:1.01–1.15); and male genitalia with the reduced, truncate apex of the anterior vesical strap and absence of the gonopore sclerite (fig. 25). Distribution of thoracic and abdominal scalelike setae as in obscurellus , rostratus , and strobicola . Distinguished from obscurellus and strobicola by the nonsexually dimorphic body and short antennal segment 2 (fig. 2), whereas these species have the male elongate and female ovate and the antennal segment 2 much greater than the width of the head (fig. 3). The fusiform antennal segment 2 and vesica with a short anterior strap (fig. 25) distinguish minusculus from rostratus , which has the antennal segment 2 of only the male slightly thickened and a long anterior vesical strap (fig. 24).
REDESCRIPTION: GENERAL ASPECT: broadly ovate; total length male 2.55 (2.33– 2.75), female 2.71 (2.50–2.90); length to cuneal fracture male 1.97 (1.88–2.20), female 2.09 (1.95–2.25); coloration dark reddish to brownish black; distal antennal segments and tarsi pale yellowish brown; claws dark; dorsum with densely distributed, flattened, broad, apically truncate and serrate, white scalelike setae and densely distributed, suberect to reclining, black simple setae; thoracic pleura and venter with scalelike setae. HEAD: width male 0.67 (0.64–0.69), female 0.69 (0.67–0.70); vertex width male 0.35 (0.34–0.36), female 0.37 (0.36–0.39); region anterior to antennal insertion elongate, anteocular length male 0.20 (0.18–0.22), female 0.20 (0.18–0.21); eyes moderately large, about 81% of head height, ventral margin of antennal insertion level with ventral margin of eye; antennal measurements male 0.18 (0.17–0.19): 0.62 (0.58–0.65): 0.39 (0.36– 0.44): 0.29 (0.28–0.31), female 0.18 (0.17– 0.19): 0.63 (0.59–0.67): 0.41 (0.38–0.44): 0.29 (0.27–0.30); antennal segment 2 is 6% shorter than head width across eyes, slightly fusiform in both sexes; labium reaching apex of middle coxae in both sexes; labial length male 1.05 (1.00–1.09), female 1.07 (1.03– 1.14); length of segment 4 male 0.30 (0.28– 0.32), female 0.31 (0.29–0.31). THORAX: width male 1.04 (0.98–1.10), female 1.06 (0.98–1.09); length male 0.53 (0.48–0.56), female 0.53 (0.50–0.56); mesoscutum relatively narrowly exposed. HEMELYTRA: maximum width male 1.34 (1.20–1.43), female 1.39 (1.30–1.50); lateral margin broadly arcuate.
BIOLOGY: Unlike other members of the genus, P. minusculus is found on species of Rosaceae rather than on conifers. Presumed hosts include apple, ( Malus sylvestris Mill. ) and hawthorn ( Crataegus chrysocarpa Ashe ). Kelton (1983) also reported it as predaceous on aphids and mites. Bouchard et al. (1988) reported the feeding voracity of P. minusculus on apple aphid ( Aphis pomi DeGeer ( Aphididae )) in southwestern Québec.
DISTRIBUTION: Eastern Nearctic ranging from Québec and Ontario in the north, south to Massachusetts, New York, and South Carolina and west to Iowa.
DISCUSSION: Knight (1923, 1931) described L. minusculus and A. crataegi from White Plains, New York and Ames, Iowa, respectively. After examining paratypes of both nominal species we must conclude that crataegi is a junior synonym of minusculus . The original descriptions of both species are practically identical. Discrepancy is found only in the measurements; however, the range of measurement in our concept of minusculus subsumes the minute differences between the original descriptions. Examination of the vesicae of both nominal species also revealed no substantive structural differences.
SPECIMENS EXAMINED: CANADA: Ontario: Aldershot, 7.VII.1955, L. A. Kelton, 1♀ ( CNC); Exeter , 12.VII.1962, Kelton, Thorpe, Crataegus sp. , 33, 2♀ ( CNC); Fort Erie , 10.VII.1955, L. A. Kelton, 13 ( CNC); Haggersville , 9.VII.1962, Kelton, Thorpe, 6♀ ( CNC). Québec: Frelighsburg , 13.VII.1977, L. A. Kelton, apple, 103, 28♀ ( CNC). USA: Connecticut: Storrs , 8.VI .1977 , D. Leston, 13 (AMNH). Iowa: Polk Co.: Mitchellville, Thomas Mitchell County Park , 29.VI.1980, K., R. Schmidt, 13 ( AMNH). Story Co.: Ames , various dates (see Knight 1931) , 21.VI .1964 , H. H. Knight, including Atractotomus crataegi paratypes, 213, 13♀ (CNC, USNM). Massachusetts: Essex Co.: Rockport, Golf Club Road , 7.VII.1999, M. D. Schwartz, Malus sp. , 13, 7♀ ( CNC). Michigan: Midland Co .: 28.VI.1958, R. R. Dreisbach, 1♀ ( USNM). New York: Albany Co.: Rensselaerville, Huyck Preserve , 29.VI– 2.VII.1977, R. T. Schuh, Malus sp. 73, 1♀ ( AMNH). Nassau Co.: Flower Hill, nr Rte 25A, Ridge Drive East, M. D. Schwartz, Malus sp. , 23, 4♀ ( AMNH). Queens Co. : Corona Heights , jct Van Doren St , 108th St ., 13.VI .1986 , M. D. Schwartz, Malus sp. , 33 (AMNH); Flushing Meadow Corona Park , near zoo, 13.VII.1985, M. D. Schwartz, Malus sp. , 2♀ ( AMNH). Westchester Co.: White Plains : 10.VII.1915, apple, Lepidopsallus minusculus paratypes, 23, 1♀ ( CNC) ; 29.VI .1929 , C. E. Olsen, 1♀ (USNM). Pennsylvania: Centre Co.: State College , 6.VI .1977 . Schuh, Henry, Wheeler, Crataegus sp. , 73, 17♀ (AMNH). South Carolina: Pickens Co.: Clemson College , 22.V .1944 , D. Dunavan, apple, 33 (CUCC). Tennessee: Claiborne Co.: Rte 25E, N of Bean Station, 21.V .1979 , Schuh, Wygodzinsky, Schmidt, 13 (AMNH). Vermont: Windsor Co.: Mount Ascutney , 23.VII.1912, Parshley, 13 ( CAS) .
Phoenicocoris modestus (MeyerDür)
Figures 1L, 2 View Fig , 13 View Fig , 23 View Fig
Capsus modestus MeyerDür, 1843: 69 View in CoL (n.sp.).
Capsus gracilicornis Scholtz, 1847: 139 View in CoL (n.sp.); Fieber, 1861: 394 (syn.).
Capsus atropurpureus Kirschbaum, 1856: 262 View in CoL , 338 (n.sp.); Baerensprung, 1860: 17 (syn.).
Sthenarus modestus form purpurascens Stichel, 1956: 366 View in CoL (new form).
Phoenicocoris modestus: Kerzhner, 1962: 231 View in CoL (n.comb.); Schuh, 1995: 375 (catalog); Kerzhner and Josifov, 1999: 387 (catalog).
DIAGNOSIS: Recognized by the relatively small size ( 2.15–2.75 mm); scutellum and hemelytra with adpressed, narrowly ovate, apically pointed, scalelike setae (figs. 1L, 13A); thoracic pleura and abdominal sternum without scalelike setae; completely pale antenna, fore and middle tibiae; labium reaching to or just beyond apices of hind coxae; and by the comparatively small vesica with a large secondary gonopore (fig. 23). The distribution of the pronotal scalelike setae in this Old World taxon is similar to those of the western North American species nevadensis and ponderosae . Distinguished from nevadensis by the short labium, which, at most, just surpasses the hind coxae, whereas in nevadensis the labium reaches the middle of the abdominal sternum. Overall body length will separate ponderosae from modestus ; in the former, body length is greater than 3.0 mm, whereas in modestus the body is less than 2.75 mm.
REDESCRIPTION: ( N = 9♀). GENERAL ASPECT: oblongate; total length male 2.46 (2.15–2.60), female 2.51 (2.30–2.75); length to cuneal fracture male 1.75 (1.63–1.93), female 1.83 (1.70–1.95); coloration reddish brown, dark reddish brown to dark brown; antennae, apices of femora, tibiae, and tarsi pale yellow; femora concolorous with dorsum; hemelytra with moderately to densely distributed, broad, flattened, apically pointed, white scalelike setae and densely distributed, suberect to reclining, pale brown simple setae; vertex, pronotum, thoracic pleura, and venter without scalelike setae. HEAD: width male 0.63 (0.61–0.64), female 0.63 (0.60– 0.65); vertex width male 0.29 (0.28–0.30), female 0.30 (0.29–0.33); region anterior to antennal insertion moderately elongate, anteocular length male 0.20 (0.18–0.21), female 0.19 (0.17–0.21); eyes moderately large, about 81% of head height, ventral margin of antennal insertion level with ventral margin of eye; antennal measurements male 0.16 (0.15–0.17): 0.57 (0.56–0.60): 0.35 (0.31–0.37): 0.27 (0.24–0.29), female 0.16 (0.15–0.17): 0.57 (0.55–0.61): 0.36 (0.31– 0.41): 0.29 (0.25–0.30); segment 2 subequal to, equal to, head width across eyes; labium surpassing apices of hind coxae in both sexes, usually reaching base of abdominal sternum; labial length male 1.27 (1.25–1.29), female 1.25 (1.23–1.26); length of segment 4 male 0.34 (0.33–0.35), female 0.37 (0.36– 0.38). THORAX: width male 0.90 (0.86– 0.93), female 0.91 (0.85–0.94); length male 0.41 (0.39–0.42), female 0.41 (0.39–0.43); mesoscutum moderately exposed. HEMEL YTRA: maximum width male 1.15 (1.05– 1.21), female 1.16 (1.08–1.20); lateral margin slightly arcuate.
BIOLOGY: Stichel (1956) recorded Scots pine ( Pinus sylvestris Linnaeus ) as a Palearctic host. All specimens housed in the Zoological Institute, St. Petersburg, Russia, with documented hosts, were collected from P. sylvestris (Konstantinov, personal commun.). Wagner (1975) noted that modestus feeds on Pinus sp. with immatures present in June and July and eggs, the diapausing life stage.
DISTRIBUTION: Widely distributed in the Palearctic Region from the Russian Far East ( Kerzhner, 1988a) south to Mongolia, west in the forest zone of Russia, Scandinavia, and Western Europe to as far west as Spain ( Kerzhner and Josifov, 1999).
SPECIMENS EXAMINED: FINLAND: Western: Parainen [Pargas], Sthenarus modestus det. by O. M. Reuter, 23, 1♀ (CNC); GERMANY: Bavaria: Mühlstetten bei Pleinfeld, Mfr, 18.VI .1988 , G. Schuster, 13 (SCHU). [Northern Bavaria]. Meyer Dür Collection, 43, 5♀ (AMNH). MONGOLIA: [in Cyrillic characters] Hantau Aimak [= district], 8 km N of Binder, 5.VII.1976, I. M. Kerzhner, 13, 2♀ ( AMNH). SPAIN: Bergueda´ : Ceres , 29.VI .1987 , J. Ribes, Pinus sylvestris , 13, 2♀ (RIBE).
Phoenicocoris nevadensis , new species
Figures 1T, 2 View Fig , 14 View Fig , 23 View Fig
HOLOTYPE: Male, ‘‘ USA: NEVADA: Churchill [Co.], reststop W. of Carroll Summit, R 37E T 16N, elev. 6500 ft., Aug. 5, 1982, coll. M. D. Schwartz, Pinus monophylla [(Torrey et Frémont) ( Pinaceae )].’’ Deposited in the American Museum of Natural History.
DIAGNOSIS: Similar to modestus in body size, color of the dorsum, antennae, and legs, and absence of scalelike setae on vertex and pronotum, but easily distinguished by the longer labium, gonopore sclerite longer than secondary gonopore, short anterior apex of the vesical strap (fig. 23), and geographical distribution. Distinguished from ponderosae , the only other species lacking scalelike setae on the pronotum, by the smaller size and vesical structure.
DESCRIPTION: GENERAL ASPECT: small; total length male 2.40 (2.26–2.60), female 2.33 (2.05–2.65); length to cuneal fracture male 1.72 (1.56–1.85), female 1.67 (1.46– 1.88); coloration dark brown to nearly black; antennae, apices of femora, tibiae, and tarsi pale yellow; antennal segment 1 with basal and distal red marks; apices of tarsal segment 3 and claws darker brown; mesoscutum, scutellum, and hemelytra dorsum with moderately to densely distributed, broad, flattened, apically serrate, white scalelike setae and densely distributed, suberect to reclining, brown simple setae; thoracic pleura and venter without scalelike setae. HEAD: width male 0.65 (0.63–0.68), female 0.64 (0.61– 0.66); vertex width male 0.30 (0.29–0.30), female 0.31 (0.30–0.33); region anterior to antennal insertion elongate, anteocular length male 0.21 (0.20–0.23), female 0.21 (0.19– 0.22); eyes moderately large, about 82% of head height, ventral margin of antennal insertion level with ventral margin of eye; antennal measurements male 0.14 (0.13–0.15): 0.50 (0.45–0.54): 0.31 (0.25–0.39): 0.26 (0.25–0.29), female 0.14 (0.13–0.14): 0.47 (0.42–0.53): 0.30 (0.28–0.33): 0.25 (0.23– 0.29); labium brown, reaching seventh sternite to base of genital segment in males and from base to middle of ovipositor in females; labial length male 1.22 (1.08–1.38), female 1.19 (1.11–1.25); length of segment 4 male 0.36 (0.33–0.38), female 0.35 (0.33–0.38). THORAX: width male 0.86 (0.84–0.91), female 0.85 (0.79–0.89); length male 0.36 (0.34–0.38), female 0.36 (0.34–0.38); mesoscutum moderately exposed. HEMELY TRA: maximum width male 1.05 (0.98– 1.10), female 1.09 (0.98–1.15); gently curved laterally.
BIOLOGY: Hosts include knobcone pine ( Pinus attenuata Lemmon ), lodgepole pine ( P. contorta Douglas ex Loudon ), piñon ( P. edulis Engelmann ), and singleleaf piñon ( P. monophylla Torrey et Frémont ).
DISTRIBUTION: Western United States from Hood River Co., Oregon in the north, south to northern and Santa Cruz Co. , California, and east to central Nevada.
DISCUSSION: We have identified a female, in poor condition, from the Hualapai Mountains of Mohave County, Arizona as our new species. We support this placement based on the small black body; short, white, scalelike setae on the hemelytra; labium reaching the base of the ovipositor; and the long parempodia with a spatulate apex. It is not unusual for plant bugs to be found on the flowers of nonhost plants, especially in this case, when the true host was present in abundance in the immediate vicinity.
PARATYPES: USA: California: Santa Cruz Co. : Mount Hermon, 8.VII. 1922, F. E. Blasidell, 1♀ (CAS). Siskiyou Co.: McCloud, Pinus attenuata : 1090 m, 7.VII.1979, R. T., J. Schuh, 33, 6♀ (AMNH); 1100 m, 9.VII.1980, R. T. Schuh, G. M. Stonedahl, 103, 2♀ (AMNH); just NW of, 3700 ft, 27.VII.1986, R. T. Schuh, 1♀ (AMNH). Nevada: Churchill Co.: rest stop W of Carroll Summit, R37E T16N, 6500 ft, 5.VIII.1982, M. D. Schwartz, P. monophylla , 63, 20♀ (AMNH). Clark Co.: Charleston Peak, J. T. Polhemus, 20.VII.1982, P. edulis , 2♀ (JTP); no locality, 29.VII.1967, D. C., K. A. Rentz, 1♀ (UCB). Lyon Co.: N boundary Toiyabe National Forest, on Rte 22, 1780 m, 11.VII.1980, R. T. Schuh, G. M. Stonedahl, P. monophylla , 113, 40♀ (AMNH). Douglas Co.: Topaz Lake, 11.VII.1967, W. J. Turner, 1♀ (UCB). White Pine Co.: 4.2 mi W of Baker, 2031 m, 14.VII.1980, R. T. Schuh, G. M. Stonedahl, P. monophylla , 16♀ (AMNH). Lehman Cave National Monument, 24.VI.1966, W. Gagne: P. monophylla , 13, 1♀ (USNM); Lehman Creek, nr, with J. Haddock, 63, 4♀ (UCB). Oregon: Grant Co. : Dixie Campground on Rte 26, Malheur National Forest, 24.VIII.1981, G. M. Stonedahl, P. contorta , 1♀ (OSU). Hood River Co.: no locality, 23.VIII.1978, J. D. Lattin, P. contorta , 2♀ (OSU).
ADDITIONAL SPECIMEN: USA: Arizona: Mohave Co.: Hualapai Mountains , SE of Kingman, T20 N R15 W, 4000–6400 ft, 9,10. VI.1983, R. T . Schuh, M. D. Schwartz, G. M. Stonedahl , Quercus sp. , 1♀ ( AMNH) .
Phoenicocoris obscurellus (Fallén)
Figures 1M, 3 View Fig , 15 View Fig , 23 View Fig
Phytocoris obscurellus Fallén, 1829: 108 View in CoL (n.sp.).
Capsus oculatus Kirschbaum, 1856: 250 View in CoL , 318 (n.sp.); Wagner, 1939: 47 (syn.).
? Atractotomus albipes Fieber, 1858: 334 View in CoL (n.sp.); Fieber, 1861: 296, (syn. with Atractotomus oculatus ).
Capsus pityophilus Flor, 1860: 463 View in CoL , 597 (n.sp.); Thomson, 1871: 446 (syn.).
Agalliastes meyeri Fieber, 1864: 231 View in CoL (n.sp.); Puton, 1873: 25 (syn.).
Atractotomus pini Fieber, 1864: 224 View in CoL (n.sp.); Douglas and Scott, 1868: 268 (syn. with C. pityophilus View in CoL ).
Psallus obscurellus form griseofusca Stichel, 1933: 262 View in CoL , 264.
Phoenicocoris obscurellus: Kerzhner, 1962: 231 View in CoL (n.comb.); Schuh, 1995: 375 (catalog); Kerzhner and Josifov, 1999: 388 (catalog).
DIAGNOSIS: Distinguished by the sexually dimorphic eyes and hemelytra; male with large eyes, elongate body, and parallelsided lateral margin of corium and female with small eyes, shorter body, and gently curved lateral margin of corium; narrow, lanceolate scalelike setae on the dorsum, thoracic pleura, and lateral margins of abdominal sternum; length of antennal segment 2 much greater than width of head across eyes; legs and antennae mostly darkened; and the gonopore sclerite nearly twice as long as the secondary gonopore (fig. 23). The sexually dimorphic body form is also found in strobicola , but the pale appendages of this North American species will distinguished it from obscurellus , which has darker legs and antenna and has an Old World distribution.
REDESCRIPTION: GENERAL ASPECT: moderately elongate (female) to elongate (male); total length male 3.52 (3.35–3.70), female 2.97 (2.80–3.20); length to cuneal fracture male 2.36 (2.23–2.48), female 2.15 (2.03–2.33); coloration dark brown to nearly black; antennae, apices of femora, tibiae, and tarsi dusky brownish yellow to yellowish brown; femora yellowish or pale brown to brown; dorsum with densely distributed, relatively narrow, flattened, apically pointed, silvery scalelike setae and densely distribut ed, suberect to reclining, black simple setae; thoracic pleura and venter with scalelike setae. HEAD: width male 0.68 (0.65–0.70), female 0.65 (0.63–0.66); vertex width male 0.25 (0.25–0.27), female 0.29 (0.28–0.30); region anterior to antennal insertion relatively elongate, anteocular length male 0.19 (0.19–0.20), female 0.20 (0.18–0.21); eyes large, 89% of head height, ventral margin of antennal insertion dorsal to ventral margin of eye; antennal measurements male 0.19 (0.18–0.20): 0.98 (0.94–1.04): 0.50 (0.48– 0.53): 0.31 (0.29–0.33), female 0.19 (0.18– 0.20): 0.83 (0.78–0.85): 0.46 (0.44–0.50): 0.32 (0.30–0.34); segment 2 is 25% to 50% longer than head width across eyes; labium brown, reaching seventh sternite in male and sternite 5 in female; labial length male 1.40 (1.26–1.53), female 1.43 (1.39–1.48); length of segment 4 male 0.38 (0.38–0.40), female 0.39 (0.38–0.40). THORAX: width male 1.00 (0.94–1.03), female 0.97 (0.93–1.00); length male 0.48 (0.44–0.51), female 0.46 (0.44–0.48); mesoscutum broadly exposed. HEMELYTRA: maximum width male 1.30 (1.25–1.40), female 1.28 (1.23–1.33); lateral margin subparallel.
BIOLOGY: Stichel (1956) listed the following conifers as hosts: Pinus sylvestris and Norway spruce ( Picea abies (Linnaeus) Karsten [as P. excelsa Link ]) ( Pinaceae ), and common juniper ( Juniperus communis Linnaeus ) ( Cupressaceae ). All specimens, with documented hosts, housed in the Zoological Institute, St. Petersburg, Russia were collect ed from P. sylvestris (Konstantinov, personal commun.).
DISTRIBUTION: Widely distributed in the Palearctic Region from the Russian Far East ( Kerzhner, 1988a) south to Mongolia, west in the forest zone of Russia, Scandinavia, Western Europe, including Great Britain and Italy, to as far west as Portugal ( Kerzhner and Josifov, 1999).
DISCUSSION: The coloration of specimens from Europe and western Russia conform to the redescription. Specimens from Yakutia and Mongolia are uniformly paler, with the dorsum and appendages pale brown or even dirty yellowish brown. Kerzhner and Josifov (1999) noted that Wagner (1939: 47) questioned the synonymy of albipes with oculatus proposed by Fieber (1861). In Wagner’s view oculatus sensu Fieber, 1861 is identical with Atractotomus magnicornis and only oculatus Kirschbaum is a junior subjective synonym of obscurellus . The synonymy of albipes with obscurellus needs to be confirmed to eliminate the possibility that albipes is rather a junior synonym of modestus .
SPECIMENS EXAMINED: ENGLAND: Bedfordshire Co.: Haynes, 16.VII.1975, D. Leston, Pinus sp. , 4♀ (AMNH); Rowney Warren, 28.VII.1975, D. Leston, 1♀ (AMNH). Hampshire Co.: Aldershot, 2.5 mi NW of, on Rte 323, 2.VII.1989, G. M. Stonedahl, Pinus sp. , 73, 23♀ (AMNH). Surrey Co.: Frensham Common, 29.VI.1994, G. M. Stonedahl, Pinus sp. , 153, 12♀ (AMNH). FIN LAND: Parainen [Pargas], Psallus obscurellus det. by O. M. Reuter, 13 (CNC); Tvärminne, H. Lindberg, 2♀ (CNC). GERMANY: Bavaria: Schwaben: Klosterlechfeld, 3.VII.1966, Gerh. Schuster, Pinus sp. , 13 (SCHU). Saxony: Oberlausitz: Creba, 6.VIII.1942, Jordan, 1♀ (AMNH); Niederschlesien, VI.1943, Jordan, 13 (AMNH); Weisskelm, 11.VII.1942, Jordan, 1♀ (AMNH). MONGOLIA: [in Cyrillic characters] Eastern Aimak, 20 km ESE of BayanUla, 8.VII.1976, I. M. Kerzhner, 33 (AMNH). RUSSIA: [in Cyrillic characters] Serezhino, Yamburgsk[iy] u[ezd] [uezd = district], 24, 25.VI.1896, Bianchi, 2♀ (AMNH). SPAIN: Osona: Collsuspina, 10.VII.1977, J. Ribes, Pinus sylvestris , 13, 1♀ (RIBE). Soria: Urbión, 11.VIII.1978, J. Ribes, 33 (RIBE).



| USNM |
Smithsonian Institution, National Museum of Natural History |
| CNC |
Canadian National Collection of Insects, Arachnids, and Nematodes |
| AMNH |
American Museum of Natural History |
| CAS |
California Academy of Sciences |
| R |
Departamento de Geologia, Universidad de Chile |
| T |
Tavera, Department of Geology and Geophysics |
| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Atractotomus Fieber, 1858 ( Stonedahl,
| SCHWARTZ, MICHAEL D. & STONEDAHL, GARY M. 2004 |
Phoenicocoris australis : Stonedahl, 1990: 60
| Stonedahl, G. M. 1990: 60 |
Phoenicocoris claricornis : Stonedahl, 1990: 60
| Stonedahl, G. M. 1990: 60 |
Phoenicocoris longirostris : Stonedahl, 1990: 61
| Stonedahl, G. M. 1990: 61 |
Phoenicocoris minusculus : Stonedahl, 1990: 61
| Stonedahl, G. M. 1990: 61 |
Phoenicocoris crataegi : Stonedahl, 1990: 61
| Stonedahl, G. M. 1990: 61 |
Lepidopsallus longirostris
| Knight, H. H. 1968: 54 |
Phoenicocoris modestus : Kerzhner, 1962: 231
| Kerzhner, I. M. & M. Josifov 1999: 387 |
| Kerzhner, I. M. 1962: 231 |
Phoenicocoris obscurellus : Kerzhner, 1962: 231
| Kerzhner, I. M. & M. Josifov 1999: 388 |
| Kerzhner, I. M. 1962: 231 |
Stenopsallus
| Kerzhner, I. M. 1962: 381 |
| Wagner, E. 1960: 12 |
Sthenarus modestus form purpurascens
| Stichel, W. 1956: 366 |
Psallus obscurellus form griseofusca
| Stichel, W. 1933: 262 |
Atractotomus crataegi
| Knight, H. H. 1931: 37 |
Lepidopsallus australis
| Blatchley, W. S. 1926: 953 |
Lepidopsallus claricornis
| Knight, H. H. 1923: 471 |
Lepidopsallus minusculus
| Knight, H. H. 1923: 472 |
Phoenicocoris : Reuter, 1876: 90
| Maw, H. E. L. & R. G. Foottit & K. G. A. Hamilton 2000: 123 |
| Kerzhner, I. M. & M. Josifov 1999: 387 |
| Kerzhner, I. M. 1962: 231 |
| Reuter, O. M. 1876: 90 |
Plagiognathus ( Phoenicocoris )
| Reuter, O. M. 1875: 99 |
Agalliastes meyeri
| Puton, A. 1873: 25 |
| Fieber, F. X. 1864: 231 |
Atractotomus pini
| Douglas, J. W. & J. Scott 1868: 268 |
| Fieber, F. X. 1864: 224 |
Capsus pityophilus
| Thomson, C. G. 1871: 446 |
| Flor, G. 1860: 463 |
Atractotomus albipes
| Fieber, F. X. 1861: 296 |
| Fieber, F. X. 1858: 334 |
Capsus atropurpureus
| Baerensprung, F. von 1860: 17 |
| Kirschbaum, C. L. 1856: 262 |
Capsus oculatus
| Wagner, E. 1939: 47 |
| Kirschbaum, C. L. 1856: 250 |
Capsus gracilicornis
| Fieber, F. X. 1861: 394 |
| Scholtz, H. 1847: 139 |
Phytocoris obscurellus Fallén, 1829: 108
| Fallen, C. F. 1829: 108 |