
Latastia ornata, Monard, 1940
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5296.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:9347E5CC-A9F3-4701-B6E0-B6D6069FAA22 |
|
DOI |
https://doi.org/10.5281/zenodo.7990576 |
|
persistent identifier |
https://treatment.plazi.org/id/3E6B87BF-0913-7675-EFA9-F8C3FD35FEBE |
|
treatment provided by |
Plazi |
|
scientific name |
Latastia ornata |
| status |
|
Redescription of external morphology
Based on a re-examination of the holotype and on the examination of the recently collected specimens RBINS 20301–20302, shown on Figures 3–6 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 . Raw measurements and scale counts of the three specimens are provided in Table 2 View TABLE 2 .
Snout-vent length to 76 mm; total length> 229 mm (holotype). Tail 2.5 times SVL (based on RBINS 20302, the only specimen with a complete, original tail; the tail tip of RBINS 20301 is missing and is not healed). Body moderately depressed. Head distinct from neck, narrow (HL/HW ratio 1.8–2.1), long (HL/SVL ratio 0.25–0.30, proportionally longest in the subadult), depressed (HH/HL ratio 0.41–0.49, least depressed in the subadult).
Head covered with symmetric plates. Rostral well visible in dorsal view. Frontonasal rounded anteriorly, slightly wider than long. A line of small tubercles along the posterior borders of the internasals, frontonasal and prefrontals (also along the lateral borders of the prefrontals in the subadult), “en forme de perles” (i.e., pearl-shaped) as described by Monard (1940). Suture between internasals subequal to suture between prefrontals (respectively 0.8 and 0.9 mm in RBINS 16301). Pupil round. Lower eyelid scaly. Canthus rostralis rounded. Lores near-vertical. Nostril opening in contact with 1 st supralabial, the anterior nasal and the two posterior, superposed, nasals. Four supraorbital scales, the anterior- and posteriormost small, separated from the supraciliaries by a continuous row of granular scales. In the holotype the anterior- and posteriormost supraoculars are entire, while in RBINS 20301 the anterior supraoculars are divided into two (left) or three (right) fragments and the posterior ones into three (left) or two (right) fragments. In RBINS 20302 the anterior supraoculars are divided into two fragments on each side, and the posterior supraoculars are unfragmented. Tympanic opening large, rounded, surrounded by smooth scales (i.e., no auricular denticulation), including a crescentic scale bordering the antero-dorsal limit of the tympanum. Seven or eight supralabials; one of them much enlarged and bordering the orbit, separating the five anterior supralabials (sometimes four, as on the left side of the holotype) from the two posterior ones. Frontal hexagonal, surrounded anteriorly by the two prefrontals, laterally by three supraoculars on each side, and posteriorly by the two frontoparietals. Anterior border of frontal pointing forward. Frontal narrow posteriorly. Length of frontal slightly smaller than distance between frontal and snout tip (respectively 4.1 and 5.1 mm in RBINS 16301). Frontoparietals pentagonal. Parietals large, nearly as long as frontal, separated by the interparietal scale and a small occipital. Pineal eye visible through the interparietal scale. Scales on the upper surface of the head smooth, except the pearl-shaped tubercles. Temporal area covered by an elongate, thin temporal plate along the parietal, and by smaller to granular scales. Six (in a single case seven) infralabials. Mental followed by four pairs of sublinguals, the first three in contact on the midline, the fourth pair separated from each other. Sublinguals progressively increasing in size posteriorly. Gular collar present and distinct, ventrally including five distinctly enlarged scales. The right profile and ventral view of the head, which had never been illustrated for the holotype so far, are shown on Figure 3 View FIGURE 3 .
Mediodorsal scale rows not widened. Dorsal scales granular, in 67–70 longitudinal rows at midbody, those on the lower flanks nearly smooth, those on upper flanks and dorsum with a median, single, longitudinal keel. About 40 dorsal scales between legs. Six longitudinal rows of parallelepipedal or trapezoidal, widened, smooth, ventral scales; the two medioventral rows narrower than the lateral ones. Between the gular collar and the line of porebearing scales, 27 or 28 transversal rows of ventrals. A distinctly enlarged preanal plate, bordered laterally by a row of a few small preanals on each side. Femoral pores in a continuous row of 17–20. The left and right series of pores of RBINS 20301 and RBINS 20302 are separated by respectively three and two poreless scales (two in the holotype according to the Figure 3 View FIGURE 3 in the original description, but actually three according to our observations). Subdigital lamellae of fingers and toes with two keels each. First finger shortest. Second finger longer than 5 th. Third and 4 th fingers longest, of subequal length. RBINS 20301 and RBINS 20302 both show 16/16 subdigital lamellae under the 4 th finger. When the leg is stretched alongside the body it extends anteriorly beyond the gular collar. Length of the feet comparable to head length. Toes without lateral denticulate fringes. First toe shortest. Second and 5 th toe of subequal length, shorter than the 3 rd and much shorter than the 4th. Subdigital lamellae under 4 th toe 23 to 26 (23 in the holotype, 25 or 26 in the two other specimens).
Supracaudals much larger than dorsal scales. All supracaudal scales rectangular, presenting a strong median keel, each keel bearing a small tubercle at its posterior extremity. Subcaudals at the base of the tail smooth and rounded posteriorly, quickly becoming rectangular with a medial strong keel, but without a terminal tubercle on the keel. The tail of the holotype is broken. Monard (1940) mentioned 87 subcaudals, but the tail tip shows a uniform color contrasting with the anterior part of the tail, indicating that it is regenerated. The tail of the subadult RBINS 20302 is original, and shows 143 subcaudals.
Coloration in life. Based on original description, RBINS 20301–20302 and Figures 3–7 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 .
The dorsal surface of the head is uniformly dark brown. The ventral surface of the head, the body and of the base of the tail is uniformly white. The background color of the sides of the head above the mouth line is dark brown, progressively darkening posteriorly to become black as the background color of the flanks and the first half of the dorsum. There is an alignment of white spots on the upper and lower lips and another on the temporal area, in continuity with four similar, irregular alignments of white spots along the whole length of the flanks. Four continuous, parallel white dorsal stripes extend from just behind the parietals till the posterior part of the dorsum where they fade and disappear ( Figures 4–5 View FIGURE 4 View FIGURE 5 and 7 View FIGURE 7 ). In the subadult these white stripes are irregular and discontinuous ( Figure 6 View FIGURE 6 ). From half-length of the trunk, the dorsal background color turns to reddish-brown (similar to the color of laterite), continuing to the tail. The upper surfaces of the proximal parts of the arms are black with white spots, turning to reddish-brown with irregular lighter spots on the distal parts of the arms. The upper surfaces of the legs show a reddish-brown background color with irregular lighter spots and markings. The lower surface of the tail progressively reaches posteriorly the uniform reddish-brown color uniformly covering the dorsal and lateral surfaces of the tail until its tip. The lower surfaces of the arms and legs are grayish-white, the palms are reddish-brown.
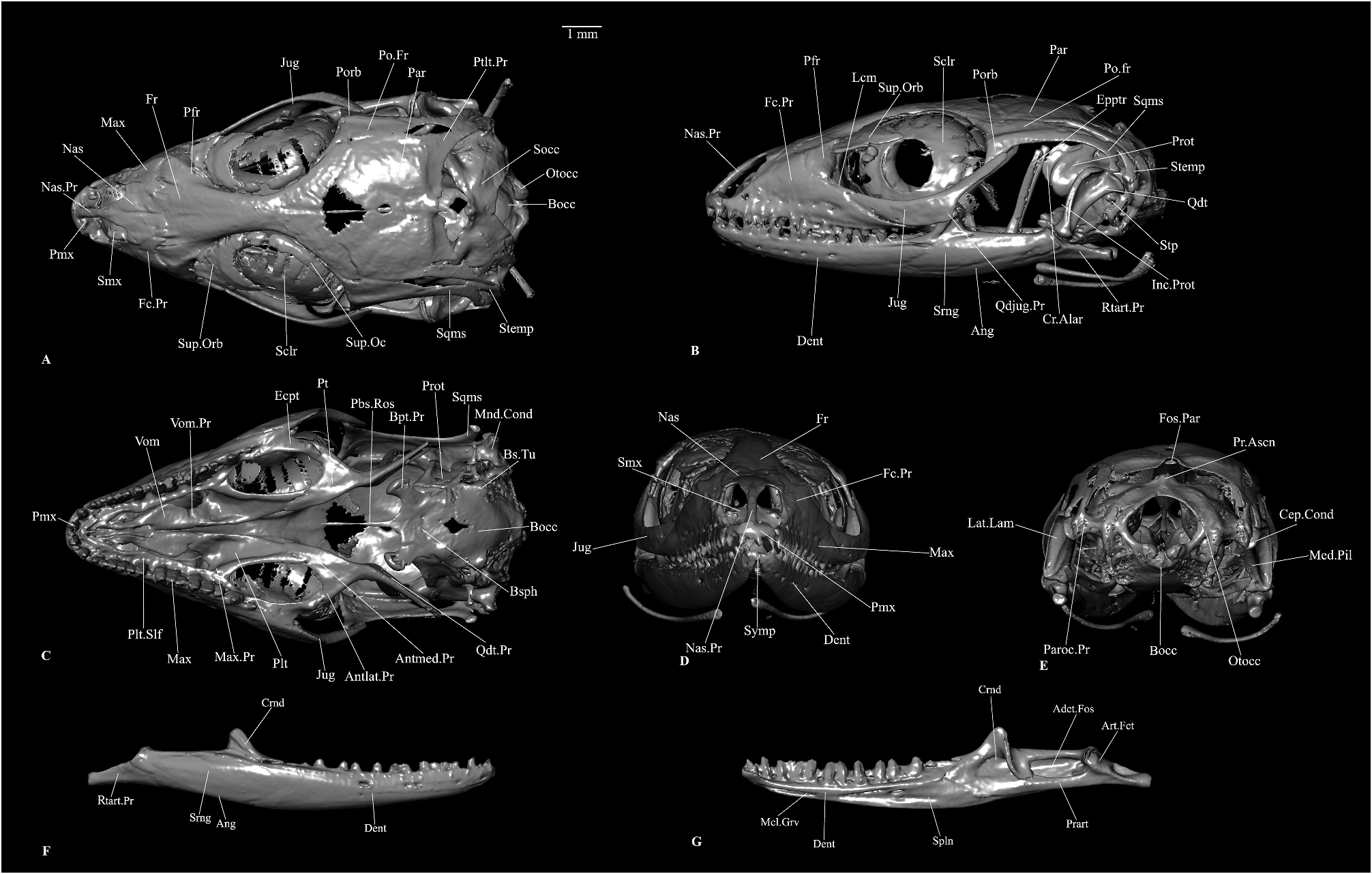
Cranial osteology. Based on subadult male RBINS 20302.
Snout and palatomaxillary bones
The premaxilla is a single, dentigerous bone ( Figure 8A–D View FIGURE 8 ). The alveolar shelf carries probably seven teeth (counting the sockets) and juts out caudad as two triangular processes touching the maxilla. There is a narrow, tapering, dorsocaudally directed nasal process that is almost as long as the nasals themselves and wedges the tapering tip between the nasals.
The nasals are paired, almost flat elements that form a straight suture between themselves and an interdigitating suture with the frontal ( Figure 8A, D View FIGURE 8 ). The anterior tips are pointed and diverge from each other to make room for the intercalating premaxillary nasal process. The nasals are the widest at about the mid-length where they project out laterally into a shark fin-shaped, anteriorly embayed protuberance slightly overlapping the maxillary facial process.
The maxilla has a high facial process with a triangular posterodorsal process reaching the frontal and another small posterolateral process below it touching the prefrontal ( Figure 8A–B View FIGURE 8 ). There is a semilunar embayment between these two processes. The alveolar border bears 15 pleurodont teeth. Right above the teeth, a palatal shelf medially expands ( Figure 8C View FIGURE 8 ). The premaxillary process is short and slightly upturned and bifurcated into an anterolateral and an anteromedial process, as is common in many lacertids ( Villa & Delfino 2019).
The palatine is an edentulous, ventrally concave bone that overlaps the palatal shelf of maxilla laterally with a maxillary process ( Figure 8C View FIGURE 8 ). This process also establishes contact with the prefrontal dorsolaterally. The squarish vomerine process overlaps the palatine process of the vomer. Between these two processes, namely the maxillary and the palatine, there is an anterior embayment and a ventral concavity corresponding to choana. Posteriorly the palatine ends in three little triangular protuberances of which the medial two overlap the pterygoid, and the lateral one laterally articulates to that bone. The palatines do not contact each other medially.
The pterygoid is an edentulous, triradiate bone ( Figure 8C View FIGURE 8 ). The anteromedial process of the pterygoid, which articulates with the palatine, is longer than the anterolateral process of the same that articulates with the ectopterygoid. The quadrate process of the pterygoid is slender, lateromedially compressed and is directed posterolateral. The dorsolateral surface of the quadrate process bears a small facet for the epipterygoid. The medial surface of this process bears a longitudinal groove for the attachment of the pterygomandibularis muscle ( Daza et al. 2011; Das & Pramanick 2019).
The ectopterygoid articulates medially to the ectopterygoid articular facet on the pterygoid anterolateral process ( Figure 8C View FIGURE 8 ). Anteriorly it overlaps the palatal shelf of the maxilla and just contacts the palatine.
The vomer forms the casing of the vomeronasal organ together with the septomaxilla ( Figure 8C View FIGURE 8 ). Vomers contact each other medially except from their posterior medial margins. Anteriorly, they contact the maxilla. Posteriorly vomers touch the palatine. Anterolaterally vomers are narrow to create an opening for the vomeronasal fenestra. Behind this, vomers expand laterally, only to taper laterally again to for choana.
The septomaxillae form the dorsal encasing of the vomeronasal organ ( Figure 8A, D View FIGURE 8 ). The septomaxillae are concave ventrally. Their medial edges are turned dorsad. Anteroventrally the septomaxilla ends with two small, pointed processes. Except a small protuberance, there is no clear posteromedial process as in some Palaearctic lacertids ( Villa & Delfino 2019).
Chondrocranial braincase bones
The braincase bones show some degree of fusion, although sutures are detectable, at least partially, between the otic capsule elements and the ventral braincase elements, namely the parabasisphenoid and the basioccipital. Prootic is a prominent element housing cochlea, anterior and horizontal (partly) semicircular canals and their ampullae ( Figure 8B–C View FIGURE 8 ). The anterior semicircular canal forms a prominent bulge on the anterolateral surface, just behind the crista alaris. In this species, the crista alaris is a narrow semilunar projection immediately rostrad and somewhat medial to the anterior semicircular canal bulge ( Figure 8B View FIGURE 8 ). Immediately below this bulge, begins a rather weakly developed crista prootica that runs caudad from this point. Ventrad to the crista prootica, the anterior margin of the prootic is embayed by the incisura prootica ( Figure 8B View FIGURE 8 ). Ventrad to the incisura prootica notch, prootic projects rostrad into an obtusely triangular anterior inferior process. The bulge along the horizontal semicircular canal continues posteriorly as a prootic process to reach the anterior surface of the paroccipital process. On the medial surface of the prootic, there are two auditory nerve foramina. The prootic articulates (in this specimen, fused) with the otoccipital along the former’s posterior lateral margin.
The paired otoccipital forms the occipital condyle with the basioccipital ( Figure 8A, E View FIGURE 8 ). The otoccipitals are composite (of opisthotic and exoccipital), hollow, bulbous bones that encase the internal ear. Dorsally the otoccipitals project out into a prominent, posterolaterally directed, axe head-shaped (in posterior view) paroccipital process that is approximately one-third the length of the quadrate. The supratemporal attaches to the anterior surface of the lateral end of this process. Ventrad to the supratemporal facet, there is a facet for the quadrate. Dorsad to the basal tubera, the otoccipital has a very prominent, deep embayment of the recessus scalae tympani which is bordered anterodorsally by the crista interfenestralis. Posterodorsal to the lateral opening of the recessus scalae tympani, there is a vagus foramen. The position of the hypoglossal foramina could not be detected clearly in the scan.
One of the two single elements of the ventral braincase is the parabasisphenoid ( Figure 8C View FIGURE 8 ). The bone has an elongated, narrow parasphenoid rostrum. Behind this parasphenoid element, the basisphenoid begins to expand. On both sides of the base of the parasphenoid rostrum, a truncated looking trabecula cranii is present. Between the trabeculae, on the dorsal surface is situated the sella turcica.Two very small internal carotid foramina open within the sella turcica. Caudad to the sella turcica, the crista sellaris is present transversely. Two short, anteroventrally directed basipterygoid processes project out from the ventrolateral base of the basisphenoid ( Figure 8C View FIGURE 8 ). These processes expand at their end. Dorsally and medially, the anterior vidian foramen pierces the basipterygoid process.
The basioccipital is the ventral element of the braincase participating in the formation of the occipital condyle ( Figure 8A, C, E View FIGURE 8 ). The bone is hexagonal, dorsally concave and in this specimen, partly fused with the otoccipital, the prootic and the parabasisphenoid. Ventrad to the lateral opening of the recessus scalae tympani, the basioccipital has a small protuberance, the basal tubera. The basioccipital forms the floor of the recessus scalae tympani.
The supraoccipital consists of a dorsomedian roof for the foramen magnum and two expanded and hollowed lateroventral wings partly encasing the inner ear ( Figure 8A View FIGURE 8 ). In this species the processus ascendens is a very small, anteriorly truncated protuberance which does not reach the parietal ( Figure 8E View FIGURE 8 ). This bone articulates with the prootic and the otoccipital.
Dermal skull roofing bones
The frontal bone is a single (fused in this specimen, condition in hatchlings is not known), elongated, skull roof element, being almost twice as long as the parietal in this species ( Figure 8A, D View FIGURE 8 ). The frontal is wide at the rostral end and very wide at the caudal end and relatively narrower in the middle. A few digitiform processes from the frontals form interdigitating sutures with the nasals and the maxillae. The frontal articulates with the prefrontal along the anterior one-third of the former’s lateral margin. On the ventral surface of the frontal, along its outer margin, there is a crest, the crista cranii. The caudal end of the frontal expands markedly into two posterolateral processes with a squarish end and articulates with the parietal and the postfrontal. This particular specimen does not show any rugosity on the dorsal surface of the frontal.
The parietal is a squarish, short, wide skull roof bone ( Figure 8A–B View FIGURE 8 ). There are no anterolateral processes. However, there is a narrow, tapering posterolateral process on each side that articulates with the supratemporal and is just separated from the squamosal. These processes do not reach the paroccipital process of the otoccipital. There is a pineal foramen ( Ledesma & Scarpetta 2018) piercing the parietal. There is a fossa parietalis ( Oelrich 1956) at the midpoint of the embayed posterior margin of the parietal, though the processus ascendens from the supraoccipital is very weakly developed in this species and does not reach the fossa.
Circumorbital bones
The prefrontals are cavernous (in anteromedial view) bones that meet the frontals laterally, the maxillary facial process anteriorly and anterodorsally, the palatines ventrally and the lacrimals ventrolaterally ( Figure 8A–B View FIGURE 8 ). The ventrolateral border of the orbitonasal flange of the parietal has a deep lacrimal notch. The orbitonasal flange projects ventrally into a posteroventral process medial to the lacrimal notch. Dorsally, the flange projects into a caudally directed process along the crista cranii. There is rather weak palpebral crest.
The jugals are paired bones consisting of an anterior and a posterodorsal process, the former being more robust, especially close to the junction between the two, than the latter ( Figure 8A–C View FIGURE 8 ). The anterior process tapers anteriorly and meets the lacrimal.This process articulates with the frontal anteroventrally and the ectopterygoid ventromedially. There is a faintly developed, caudally directed protuberance, the quadratojugal process, at the junction between the anterior and the posterodorsal processes. The slender posterodorsal process touches the postorbital.
The lacrimal is a small sliver of a bone overlying the junction of the prefrontal, jugal and the maxilla (the prefrontal is just separated from the jugal), immediately caudad to the prefrontal lacrimal notch ( Figure 8B View FIGURE 8 ).
The postfrontal is a triradiate bone with a very slender anterolateral process articulating to the posterior lateral margin of the frontal, a weakly developed, triangular protuberance articulating to the anterodorsal margin of the postorbital and a robust, dagger-like posterior process wedged between the frontal and the parietal medially and the postorbital laterally ( Figure 8A–B View FIGURE 8 ).
The postorbital is situated below the postfrontal ( Figure 8A–B View FIGURE 8 ). It consists of a short, triangular anteroventral process that contacts the posterodorsal process of the jugal and a longer posterior process touching the postfrontal and the squamosal, but the resolution of the scan did not permit determination of the exact end point of this process. In this specimen, the postfrontal and the postorbital do not extend more than one-third the length of the squamosal and thus, leave a prominent upper temporal fenestra open.
There are prominent supraorbital ossifications above and scleral ring within the orbit ( Figure 8A–B View FIGURE 8 ).
Suspensorial and palatoquadrate derived bones
The squamosals are a pair of J-shaped suspensorial bones ( Figure 8A–B View FIGURE 8 ). Rostrally the squamosal is tapered and articulates medially to the postorbital. Posteriorly the bone curves ventrad and contacts the supratemporal dorsomedially and the quadrate ventrally. The ventrally curved caudal end of this bone is squarish.
The supratemporals are small, curved bones with a tapered rostral and widened caudoventral end ( Figure 8A–B View FIGURE 8 ). The caudoventral end bears an articulatory facet for the quadrate. Anterolaterally and anteromedially the supratemporal articulates with the squamosal and the posterolateral process of the parietal, respectively.
The epipterygoid is a rather nondescript rod-like bone that articulates to the pterygoid right behind the ectopterygoid articulation. Its dorsal part closely approaches the crista alaris ( Figure 8B View FIGURE 8 ).
The quadrate consists of a medial pillar and a lateral lamina with a deep concavity on its posterior surface ( Figure 8B, E View FIGURE 8 ). There is no medial lamina. The dorsal end of the pillar expands posteromedially into a cephalic condyle that contacts the squamosal, the supratemporal and the paroccipital process ( Figure 8E View FIGURE 8 ). The ventral end of the pillar expands lateromedially into a weakly bicipital mandibular condyle that articulates with the compound bone ( Figure 8C View FIGURE 8 ). A stapes with a shaft and footplate is situated at the fenestra ovalis ( Figure 8B View FIGURE 8 ).
Mandible
The mandible is discussed here as a single element as the resolution of the scan does not permit determining the exact extent of the individual bones. The dentigerous element is the dentary ( Figure 8B, D, F, G View FIGURE 8 ). There are 20 teeth on the dentary. The posterior end of this bone is forked but the exact extents could not be determined. The anterior end of the same has a small symphysial articulatory surface ( Figure 8D View FIGURE 8 ). The medial aspect of the dentary bears a Meckelian groove ( Figure 8F View FIGURE 8 ). There is a coronoid bone, sitting on the dentary and the compound bone, with a prominent dorsally directed, obtusely triangular coronoid process, a bifurcated anterior end and a medial ridge along the posterior margin of the bone ( Figure 8F–G View FIGURE 8 ). The splenial is present but its outline could not be clearly discerned in our scan ( Figure 8G View FIGURE 8 ). The compound bone consists of the lateral surangular element, the lateroventral and ventral angular element, medial and posterior prearticular and a small articular ( Figure 8B, F, G View FIGURE 8 ). The retroarticular process probably has a heavy contribution from the prearticular, besides the articular ( Good 1987; Ledesma & Scarpetta 2018). The retroarticular processes are straight, somewhat tapering, have a faint lateral ridge and a truncated caudal end ( Figure 8B, F, G View FIGURE 8 ). Immediately rostrad to this process is an articular facet for the quadrate ( Figure 8G View FIGURE 8 ). In front of the articular facet, bounded by the surangular, the angular and the prearticular, is an adductor fossa which is visible in the dorsal and medial views ( Figure 8G View FIGURE 8 ).
Molecular analysis
Three sequences of 1143, 392 and 394 base pairs were obtained for the cytb, 12S and c-mos genes, respectively. They showed the highest similarity with sequences of Latastia longicaudata , with p-distances of 16.9%, 7.7% and 0%, respectively. For cytb and 12S, the p-distances observed between Latastia ornata and L. longicaudata (16.9–18.1% and 7.7–8.4%) were in the range of interspecific and intrageneric p-distances among the Eremiadinae analysed here (1.4–21.6% and 0.3–14.4%). For c-mos, no substitutions were observed between Latastia ornata and L. longicaudata while p-distances of 0.9–5.1% were found among species of the same genus.As one single specimen of L. ornata was sequenced, we have no information of the intraspecific variability in the species. Concerning Latastia longicaudata , intraspecific p-distances were between 0.3 and 16.8% for cytb and between 0 and 5.8% for 12S (only one sequence available for c-mos) and overlapped with the interspecific and intrageneric p-distances among the Eremiadinae analysed here. In all phylogenetic trees, the sequences of L. ornata generated here clustered with L. longicaudata ( Figures 9–11 View FIGURE 9 View FIGURE 10 View FIGURE 11 ).
Updated diagnosis
Based on males only (females unknown). A medium-sized lizard (SVL to 76 mm), with a rounded snout, slender body, well-developed limbs and a long tail (about 2.5 times SVL). Nostril opening in contact with 1 st supralabial. Five (sometimes four) anterior supralabials. Occipital plate present. Mediodorsal scales not enlarged compared to other dorsals. Gular collar present and distinct. DSR 67–70 at midbody. Six longitudinal rows of smooth ventral scales. Femoral pores in a continuous row of 17–20 on each side. Toes without lateral denticulate fringes. Maxillary teeth <20, frontal caudally twice wider than its anterior end’s width, width of the anterior process of the jugal being only twice that of the posterodorsal process of the same. Dorsum black with four continuous white lines anteriorly, turning posteriorly to reddish-brown. Throat and belly white. Tail reddish-brown.
Geographic distribution
The new localities are situated within the Prefecture of Kérouané , Kankan Region, Guinea-Conakry ( Figure 12 View FIGURE 12 ; Table 1 View TABLE 1 ). These new localities represent a major range extension of ca. 700 km towards the SE. The altitudes of the new localities vary between 629 and 858 m asl, which represents a major extension from the low altitude of 10 m asl at Bafatá, Guinea-Bissau, the type-locality.









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.