
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/10.1002/jmor.10059 |
|
DOI |
https://doi.org/10.5281/zenodo.3811871 |
|
persistent identifier |
https://treatment.plazi.org/id/3A61405D-5955-1042-FCFA-FB278A3FFC1F |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
This article develops a functional model for the arctometatarsus that incorporates inferences of soft tissues. Our format follows the logical progression involved in developing and testing a complex hypothesis of locomotion: 1) We first introduce aspects of bone and connective organ function in large terrestrial vertebrates that suggest hypotheses of arctometatarsus morphology and action. 2) Our investigations and results test these proposals and lead inductively to a model of overall arctometatarsus function. 3) Because the model is emergent from the results, we present it in a separate section. 4) The subsequent discussion tests the model against modern analogs and addresses implications of the proposed kinematics for tyrannosaurids.
The model therefore acts as a prism that focuses multiple lines of evidence into a coherent functional picture and a fulcrum over which morphological detail facilitates discussion of behavior. Our construction and application of the model exemplify the integrated approach necessary for reconstructing locomotion of extinct vertebrates.
Arctometatarsus as an Elastically Damped System
As with foot structures in modern animals, the arctometatarsus’ role in tyrannosaurid locomotion hinged on the specific morphology of its bones and connective tissues. In vertebrates, tendons that join bone to muscle, and ligaments that connect bone to bone, transmit and dissipate locomotor forces acting on the skeleton ( Hildebrand, 1988). Tendons and ligaments are variably elastic, depending on their relative proportions of collagen and elastin proteins ( Alexander, 1988) and the magnitude and period of imposed loadings ( Pollock, 1991). The bones of the arctometatarsus and its unmineralized tissues would have been elastic to some degree, but understanding its precise function depends, in part, on consideration of locomotor elasticity in modern vertebrates.
A number of studies have explicated the locomotor role of elastic ligaments and tendons and their interactions with associated bones. Elastic fore- and hindfoot connective elements store, return, and distribute footfall energies and forces. Ligaments of the feet of humans ( Kerr et al., 1987; Alexander, 1988; Deane and Davies, 1995) and the wrists of horses ( Rubeli, 1925) are noteworthy examples.
Ligaments absorb shock under specific physical conditions. Paradoxically, they display greater strength and resiliency when subject to high magnitude, sudden loadings, such as those incurred during rapid locomotion ( Frank and Shrive, 1994). In animals of large body size, the extensibility of connective elements increases because their cross sectional area is lower relative to mass than ligaments and tendons of smaller animals ( Pollock, 1991). The ligaments of large vertebrates store and return relatively more elastic strain energy, which increases locomotor efficiency and decreases strain energy transmitted to bones ( Pollock, 1991). These conditions probably existed in adult tyrannosaurids, which ranged from two ( Paul, 1988) to an upper estimate of eight tons (cross scaling of measurements from fragmentary metatarsals: University of California Museum of Paleontology, locality number V91181 View Materials ).
These considerations suggest that elastic mechanisms may have operated efficiently within the tyrannosaurid foot. Two contingencies are critical for assessing possible elastic functions: the type of connective tissues associated with the arctometatarsus and the freedom of movement between metatarsals. These factors lead to hypotheses of anatomy and movement necessary for erecting a comprehensive model of tyrannosaurid arctometatarsus function.
Prerequisites for Modeling Arctometatarsus Function: Reconstructing Soft Tissues and Intermetatarsal Displacement
Soft tissue reconstruction of extinct vertebrates entails comparison with extant relatives, ideally by bracketing the extinct taxon between modern, derived representatives of its clade and a more archaic living sister group ( Bryant and Russell, 1992; Witmer, 1995). Although there are no extant arctometatarsalian theropods, unpreserved tissues are still inferable through broader phylogenetic comparison, mechanical considerations, and universal correlates of soft parts that are present on mineralized structures ( Bryant and Seymour, 1990; Bryant and Russell, 1992).
Soft tissues leave consistent marks on bone, termed osteological correlates ( Witmer, 1995), that are evident in both extant and fossil specimens. Ligament or tendon attachments display two primary correlates: rugosity, and rough or smooth faceted areas. Rugosity marks the location of Sharpey’s fibers, mineralized collagen fibers within the bone that are continuous with fibers of the attaching connective element ( Woo et al., 1987). Ligaments and tendons also attach through a gradient of fibrocartilage, mineralized fibrocartilage, and bone. These so-called direct insertions occur on bone surfaces that are smooth and slightly concave ( Doglo-Saburoff, 1929). Sutured or closely conforming bones invariably indicate ligament attachments, and more widely spaced adjacent facets may indicate tendon attachments ( Gray and Goss, 1959).
Using these criteria we identified osteological correlates along intermetatarsal articular surfaces of large theropods. Ligaments would indicate passive elasticity and tendons would signify controlled function. Our study tested the following hypotheses: H1: Connective tissue scars on adjacent tyrannosaurid metatarsals indicate tendons. H2: Scar configuration was identical in tyrannosaurids and in the nonarctometatarsalian theropod, Allosaurus fragilis .
Freedom of movement between tyrannosaurid or Allosaurus fragilis metatarsals dictated the function of their associated ligaments or muscles. Therefore, this study sought to test hypotheses of possible intermetatarsal displacement. Preliminary observations of tyrannosaurid metatarsals, and the work of Wilson and Currie (1985) and Holtz (1995), suggested this hypothesized pattern of movement: H3: In the tyrannosaurid metatarsus, the distal end of MT III was free to move anteriorly to a small degree about a pivot point at the proximal end.
The proximal articulation between metatarsals, and their associated soft tissues, would constrain this potential displacement of MT III. Detailed consideration of both osteological and soft tissue anatomy was therefore necessary to test the hypotheses and come to a more complete understanding of tyrannosauruid arctometatarsus function.
Institutional abbreviations. MOR: Museum of the Rockies, Bozeman, Montana. IVPP: Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China. NMC: National Museum of Canada, Ottawa. PJC: Philip J. Currie, Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta, Canada. TMP: Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta. UCMP: University of California Museum of Paleontology, Berkeley, California. ROM: Royal Ontario Museum, Toronto, Ontario.
MATERIALS AND METHODS
For comparative purposes, specimens of large and small arctometatarsalian forms were examined at UCMP, MOR, and TMP. Metatarsal specimens of Allosaurus fragilis (MOR 693), and others at MOR and TMP, provided control representatives of the primitive condition for theropods. The specimens ( Table 1 View TABLE 1 ) were sufficiently complete and well preserved for evaluation of osteological correlate position and/or resolution of possible intermetatarsal movement.
Assessment of metatarsus dynamics in tyrannosaurids entailed three related lines of inquiry: 1) We identified and measured osteological correlates of soft tissues in tyrannosaurids and Allosaurus fragilis . The proximity and complexity of joint surfaces tested whether the correlates indicated tendons or ligaments. 2) To ascertain the probable range of motion between elements in physical specimens, we manipulated casts of Tyrannosaurus rex metatarsals. 3) To evaluate intermetatarsal freedom of movement in other tyrannosaurids, we examined com- puted tomographic (CT) images of Albertosaurus sarcophagus and Gorgosaurus libratus metatarsals.
Assessment of Osteological Correlates
We ascertained the distribution and extent of osteological correlates on theropod metatarsals by identifying probable scars and measuring their areas. Likely correlates of soft tissues were identified on specimens in a satisfactory state of preservation, using the criteria of rugosity and delineated faceting outlined above. Surfaces had to be continuous with cortical bone that had not been taphonomically eroded, to avoid the potential of infilled spongy bone being mistaken for rugosity. Problematic degeneration was not present on the metatarsals chosen for area measurement.
We measured the area of the correlates only when their boundaries could be repeatably determined, but because soft tissues are not preserved, our techniques are subject to some inaccuracy. Our results, therefore, reflect the position and relative area of correlates more than their absolute extent.
Surface areas of osteological correlates were measured on specimens of Albertosaurus sarcophagus (MOR 657), Allosaurus fragilis (MOR 693), Daspletosaurus torosus (MOR 590), and Tyrannosaurus rex ( MOR 555 ). The bones were wrapped in plastic cling film and attachment surface areas traced with a water-based marking pen. This technique facilitates area measurement of complexly contoured surfaces ( Snively, 2000). The cling wrap was removed from the bone, pulled gently taut, and smoothed with a ruler. The markings were then retraced onto white paper and digitized. From these scans we determined the areas of the representations in cm 2 using NIH Object-Image for Macintosh.
The average of apparent attachment areas on adjacent bones was used to approximate the cross sectional area of intervening soft tissues. A disarticulated MT IV was not present in MOR 555. To estimate the areas on that bone, first the smaller ratio was found between MT IV and MT III in the other tyrannosaurids. This ratio was then multiplied by the corresponding MT III area in MOR 555.
Manipulation of Tyrannosaurus rex Casts
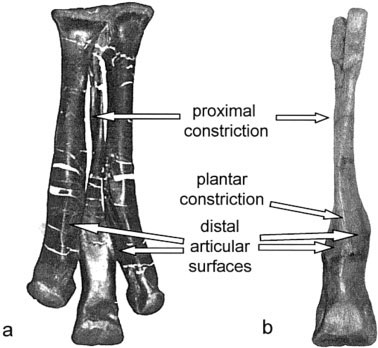
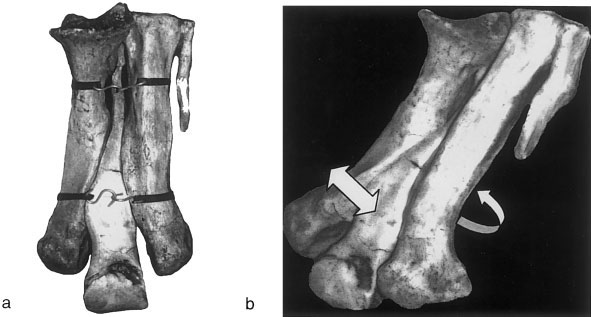
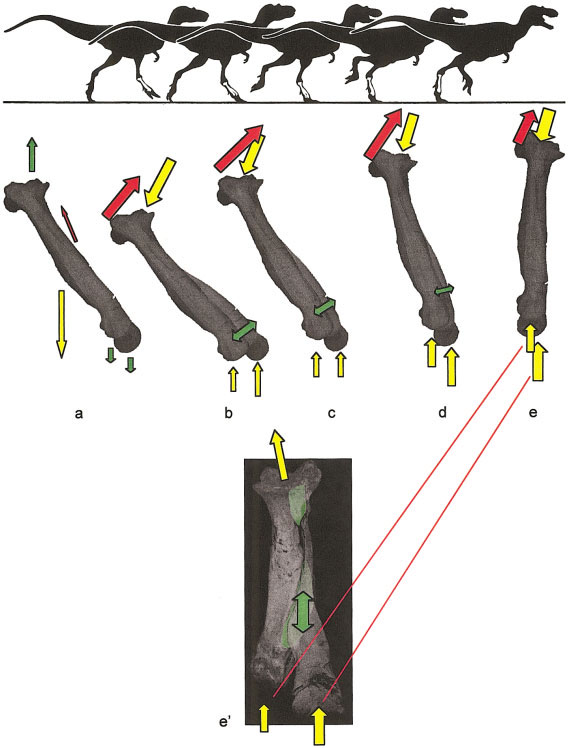
Casts of Tyrannosaurus rex metatarsals from the left foot of LACM 7244/23844 (TMP casts: 82.50.7) View Materials were positioned in proper articulation and wrapped with elastic bands. Rubber and polyester fiber bungy cords of low stiffness (0.75 meters long when untensed) were stretched and wound twice around the casts at their proximal and distal ends, tightly enough for the ends to be secured together by their plastic hooks ( Fig. 2a View Fig. 2 ).
The casts were positioned and manipulated in several ways in order to investigate proximal and distal freedom of intermetatarsal movement.
Proximal movement. We placed the distal part of the metatarsus on a laboratory bench, alternately on its anterior and posterior surfaces, while we supported and manipulated the proximal end to assess possible intermetatarsal movement in this region.
Distal movement with the metatarsus in varying positions. We placed the proximal end of the metatarsus on the bench, with the posterior surface down, and supported the specimen by the third metatarsal. Rotating the metatarsus about its fixed proximal end revealed the passive displacement of MT II and MT IV relative to MT III, with the metatarsus in various positions, ranging from 0 to –90° from the horizontal. (Some of these positions are shown in the silhouettes in Fig. 5 View Fig. 5 .)
Intermetatarsal displacement under simulated loading. The entire metatarsus was set on the bench. Taking care not to apply medial or lateral pressure, we pushed down on the dorsal surfaces of MT II and MT IV about 70% of their lengths from their proximal ends. We then lifted the distalmost portion of MT III. This showed the behavior of the outer metatarsals when a greater dorsally directed torque was applied to the distal part of MT III than to the distal ends of the other metatarsals.
CT Scanning of Tyrannosaurid Metatarsals
The methods described above apply to overall intermetatarsal movements. The topographical and likely functional complexity of the arctometatarsus compelled analysis of movement evident in cross sections at multiple transverse and longitudinal transects. As in clinical practice, the most common nondestructive technique for macroscopic paleontological sectioning is CT scanning.
In order to maximize the information from CT scanning and postprocessing visualization techniques, particular care was taken in specimen choice and preparation. TMP 94.12.602, a partial skeleton of Gorgosaurus libratus from the Dinosaur Park Formation (Late Campanian) of Dinosaur Provincial Park, Alberta, has a complete right metatarsus that has not been disarticulated taphonomically or through preparation. This specimen is undistorted and is intermediate in length and robustness between metatarsi of subadult Albertosaurus sarcophagus and adult Tyrannosaurus rex . For these reasons TMP 94.12.602 was deemed a suitable compromise for functional extrapolation to other tyrannosaurids.
The specimen was imaged at Children’s Hospital and Health Center, San Diego. Prior to scanning the specimen was encased in plasticine to a depth of 2.5–4 cm to reduce the density gradient between the bone and the surrounding medium, and thus diffraction and scattering artifacts. Once prepared, TMP 94.12.602 was CT scanned at 140 kVp and 170 mA, technique settings that produce good readings from dense bone. The cross sectional data were reconstructed in three dimensions and these reconstructions were further sectioned, viewed, and printed in various orientations for study.
Additional CT scans of an Albertosaurus sarcophagus metatarsus (TMP 81.10.1) were performed at the Radiology Department of Foothills Hospital in Calgary, Alberta. A metal frame on the specimen caused diffraction artifacts, which we alleviated by adjusting contrast on the output images. Following this adjustment the shape of intermetatarsal articulation surfaces could be evaluated and possible relative movement determined in a given plane of section.
RESULTS
Osteological Correlate Reconstruction
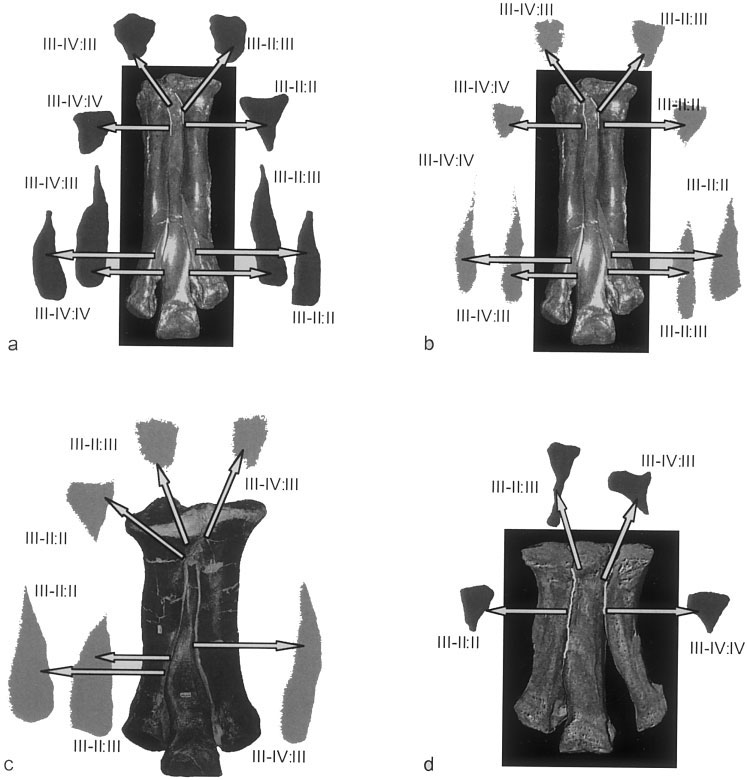
The following intermetatarsal osteological correlates for soft tissues were identified. The surfaces of all but one scar (in Allosaurus ) conform closely to the contours of the facing scar on the adjacent metatarsal. Rugosity indicating Sharpey’s fibers occurs at proximal shaft-to-shaft articular surfaces in specimens of tyrannosaurids ( Fig. 4 View Fig. 4 a–c) and Allosaurus ( Fig. 4d View Fig. 4 ). Smooth articular facets extend the MT III-MT II articulation distally in Allosaurus ( Fig. 4d View Fig. 4 ). Indications of Sharpey’s fibers are most striking along the distal articular surfaces between the shafts of MT II and MT III in large tyrannosaurids ( Fig. 4 View Fig. 4 a– c), but are not present at these locations in Allosaurus ( Fig. 4d View Fig. 4 ). A faceted distal MT III-MT IV articulation occurs in tyrannosaurids ( Fig. 4 View Fig. 4 a–c), but is entirely absent in Allosaurus , in which MT IV shows pronounced lateral angulation ( Fig. 4d View Fig. 4 ).
Proximal scars are subtriangular in all assessed theropods ( Fig. 4 View Fig. 4 ), but in Allosaurus are long proximodistally relative to the length of the metatarsus. In all three tyrannosaurids distal articulations are long and taper proximally. The average areas of adjacent distal scars are quite extensive in tyrannosaurids ( Table 2 View TABLE 2 ). The average distal scar area exceeds the average proximal area by 1.5728 in Daspletosaurus (MOR 590), 1.4029 in Albertosaurus (MOR 657), and by 1.4666 in Tyrannosaurus ( MOR 555 ; Table 2 View TABLE 2 ).
The intimate conformity of adjacent scars falsifies hypothesis H1, that tendons and muscles rather than ligaments were present between theropod metatarsals. Large distal osteological correlates on tyrannosaurid metatarsals, and their absence in Allosaurus fragilis , falsify the hypothesis (H2) that there were no marked differences in correlate morphology between the taxa.
Intermetatarsal Movement
1. Physical manipulation of Tyrannosaurus rex casts.
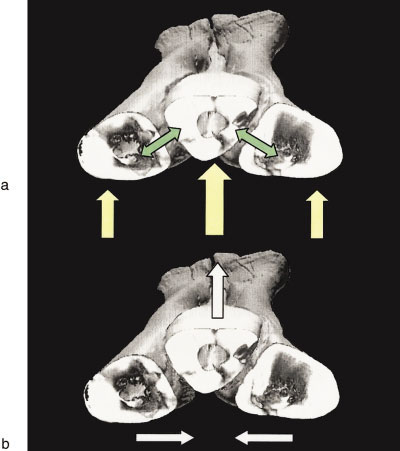
Proximal Movement. Proximally, displacement is greatly constrained by the hooked cross section of MT III and its articulation with anterolateral and posteromedial projections of MT II and MT IV, respectively ( Fig. 2b View Fig. 2 ).
Distal Movement. The distal portion of the third metatarsal is free to move anteriorly, corroborating hypothesis H3. When the anterior face of the metatarsus is parallel with the ground, only the distal elastic bands prevent this portion of MT III from pivoting towards the floor, with its center of rotation at the proximal clasped articulation.
When the posterior surface of the metatarsus faces the ground and the distal and proximal parts of MT III are fixed in position, the distal portions of MT II and MT IV slide ventrally and towards the centerline of MT III. MT II slides in a straight line along its articular surface with MT III. This contrasts with MT IV, which slides in more of an arc along its corresponding surface ( Fig 2b View Fig. 2 ).
Intermetatarsal displacement under simulated loading. When the posterior surface of the metatarsus again faces down, as just described, but with MT II and MT IV fixed proximally and distally, the same medial sliding motion occurs when the distal part of MT III is forced upwards and the bands stretch. As force is released from MT III, the bands recoil and the metatarsals return to their original articulation positions.
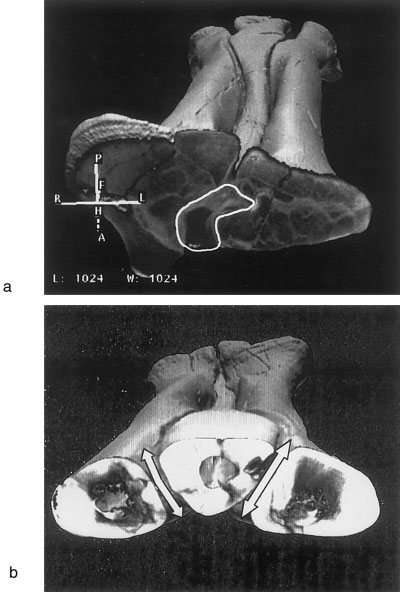
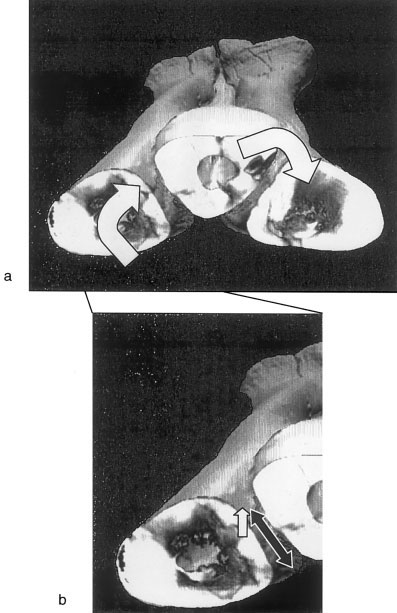
2. Freedom of movement inferred from Gorgosaurus libratus and Albertosaurus sarcophagus CT scans. These results show no gross variation in potential movement among the three metatarsi. The CT scanned specimens show the same proximal interlocking morphology as described for Tyrannosaurus rex . In Gorgosaurus libratus (TMP 94.12.602), cross sections along the metatarsus reveal that the distal articulation between MT II and MT III always slants ventromedially at the same angle ( Fig. 3a View Fig. 3 ). This indicates that displacement along this articulation will be in one
plane, a motion identical to that possible in the physical model (see results for the cast manipulations above). By contrast, the MT IV-MT III articulations in the cross sections are not always in a straight line and the overall angle of the articulation varies with cross section. This corroborates the inference that motion along this articulation would transcribe an arc in the metatarsi of all three tyrannosaurids ( Fig. 3b View Fig. 3 ).
Kinematic Model of the Tyrannosaurid Arctometatarsus: The Tensile Keystone Hypothesis
The above results indicate that ligaments were the connective elements between adjacent tyranno- saurid metatarsals and reveal degrees of freedom and constraint for tyrannosaurid intermetatarsal movement through the ground contact (stance) phase of the step cycle ( Figs. 5– 6 View Fig. 5 View Fig. 6 ). These considerations suggest the following pattern of intermetatarsal kinematics when the foot was in contact with the substrate.
1. The foot pads ventral to the phalanges would contact the substrate initially ( Fig. 5b View Fig. 5 ). Groundreaction forces would transfer to the metatarsals first across the metatarsophalangeal joints and then to the portions of the foot pad ventral to the respective metatarsals. This sequence has been corroborated through observations of domestic chickens and ostriches.
2. Because metatarsal III (MT III) is longest, the ground-reaction force would act upon the longest moment arm from the mesotarsal to the phalangeal joints. This torque differential would displace MT III anterodorsally relative to MT II and IV ( Fig. 5b,c View Fig. 5 ). Anteroposterior rotation of the proximal portion of MT III, as suggested for the small arctometatarsalian Troodon inequalis ( Wilson and Currie, 1985), was not possible in tyrannosaurids ( Fig. 3a View Fig. 3 ). Instead, the clasped proximal articulation between metatarsals would serve as a pivot point for distal rotation of MT III ( Figs. 5 View Fig. 5 b–e, 6b).
3. Crucially, forces from this differential loading and displacement pattern would stretch distal intermetatarsal ligaments ( Fig. 5b View Fig. 5 ). Upon rebound ( Fig. 5c,d View Fig. 5 ), the angulation of metatarsals, and orientation of ligaments, would draw the distal portion of the lateral metatarsals together ventrally and towards the midsagittal plane of the third metatarsal ( Fig. 6 View Fig. 6 ).
4. Forces from anterior displacement of MT III, which stretched intermetatarsal ligaments in the manner described, would decrease as the metatarsus became vertical and parallel with the ground-reaction force. In this position, ground-reaction loadings on MT III would be transferred laterally via MT II and MT IV to the condyles of the astragalus ( Wilson and Currie, 1985; Holtz, 1995). Tensile loading on intermetatarsal ligaments would mediate the energy transfer ( Fig 5 View Fig. 5 e’).
This pattern of movement has several implications. The distal arctometatarsus would become more unitary under high initial footfall loadings ( Fig. 6 View Fig. 6 ). In effect, the outer metatarsals would “splay” away from the center midline of MT III only as forces lessened, returning to their unloaded configuration.
Upon strongly oblique or torsional footfalls, ligaments and the imbricate distal cross section of the metatarsals ( Fig. 7 View Fig. 7 ) would strongly arrest interelement shear. Potentially damaging torsion of the metatarsus would be induced during abrupt turns in which torque was insufficient to overcome friction between the foot pad and the ground. The plantar angulation between metatarsals would ensure that torsional loadings were transferred from one meta- tarsal to the next ( Fig. 7a View Fig. 7 ) and would obviate anteroposterior shear. The large cross sectional area and consequent stiffness of the distal intermetatarsal ligaments ( Figs. 4 View Fig. 4 , 7b View Fig. 7 ) would check lateral shearing components introduced by torsion.
We propose the appellation “tensile keystone model” for these kinematics. Although the loading regimes are inverted, one can think of the distal part of MT III and its ligaments as analogous to the keystone of a Roman arch, in which the central element imparts stability to the entire structure.
DISCUSSION
Comparisons with Modern Analogs: Implications of Soft Tissue Scars
The results indicate that correlates of soft tissues present on large theropod metatarsals were liga- ment scars. In extant vertebrates, muscles and tendons do not normally attach on facing medial and lateral surfaces of closely joined, weightbearing elements. The closely adpressed articular surfaces of theropod metatarsals argue against the presence of muscles; negligible fiber lengths would prevent the muscles from performing positive work.
In contrast to the rarity of muscles and tendons, proximal ligaments are common between metatarsals ( Kerr et al., 1987; McGregor, 2000). Among squamates, ligaments with oblique distolateral angulation are present in the metacarpus and metatarsus of lizards ( Landsmeer, 1981; McGregor, 2000). The presence of oblique deep ligaments in lizards does not allow bracketing for homologous ligaments within the theropod metatarsus. However, it does show that ligaments of similar angulation to that hypothesized for the arctometatarsus are mechanically feasible.
The strong inference of osteological correlates as ligament scars corroborates the tensile keystone model. However, possible extant analogs can potentially falsify the hypothesis if their behavior contradicts expectations of bone–ligament function.
Comparisons With Modern Analogs: Horse Wrists and the Feet of Cursors
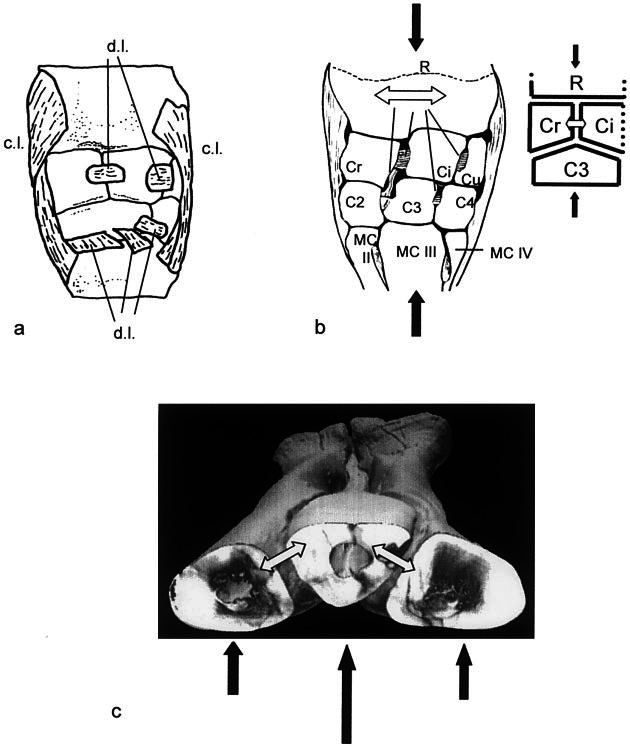
Several aspects of the tensile keystone model, and Holtz’s (1995) complementary hypothesis of energy dyles: Holtz, 1995) in a similar manner ( Fig. 8c View Fig. 8 ). Horse interosseous carpal ligaments stretch and rebound under high momentary loadings ( Fig. 8b View Fig. 8 ) ( Rubeli, 1925) and the same would be expected for tyrannosaurid intermetatarsal ligaments ( Fig. 8c View Fig. 8 ) ( Frank and Shrive, 1994).
The shear- and torsion-resisting aspects of the tensile keystone model also find analogs in the equine wrist. Wedge-like articulations generally resist shear between horse carpals ( Boening, 1981). A triangular sagittal projection of the distal radius ( Figs. 1c View Fig. 1 , 8 View Fig. 8 ) buffers ad- or abductional torsion ( Poplewski, 1936). Faces of this projection act as stop facets ( Yalden, 1971) against dorsomedial or dorsolateral rotation of the radial and intermediate carpals. Tyrannosaurid metatarsals and metapodial ligaments would function analogously by arresting torsional forces. Unlike the horse morphology, how- transference, conform remarkably with the functional morphology of the wrist (carpus) of horses. For example, the horse carpus attains high aggregate surface area between individual carpal bones, with the development of wedge-like amphiarthroses and a full complement of elements ( Figs. 8a View Fig. 8 ). High surface area decreases pressure impinging on any one carpal surface and pressure transmitted to the radius ( Bourdelle and Bressou, 1972). In the arctometatarsus distal ligaments and the distal plantar angulation of elements increased total articulation surface area and ligament cross section ( Fig. 4 View Fig. 4 ; Table 2 View TABLE 2 ), which probably conferred a similar benefit.
Rubeli (1925) demonstrated an additional advantage to the wedge-and-ligament morphology of the horse carpus, which has dorsal ligaments on the anterior surface and deep interosseous ligaments between carpals ( Fig. 8a,b View Fig. 8 ). Interosseous ligaments transduce sudden compressive loadings into a collectively longer period of elastic loadings, reducing the rate of strain. Ligaments in the arctometatarsus may have mediated the transfer of compressive forces across the ankle joint (to the astragalar con- ever, these elements would primarily buffer torsion about the midsagittal axis ( Figs. 1c View Fig. 1 , 8 View Fig. 8 ).
A more fundamental distinction between horse intercarpal and tyrannosaurid intermetatarsal ligament function lies in the initial loading regime upon footfall. The horse third metacarpal, the single weight-bearing element of the anterior metapodium, transfers compressive forces directly to the carpus ( Figs. 1c View Fig. 1 , 8 View Fig. 8 ) ( Rubeli, 1925). The carpus acts as a shock absorber for the compressive ground-reaction force. Under the tensile keystone model, dorsally directed components of the ground-reaction force load the three tyrannosaurid metatarsals unevenly ( Figs. 5 View Fig. 5 , 6 View Fig. 6 , 8c View Fig. 8 ). The third metatarsal is displaced anteriorly relative to MT II and MT IV; differential forces stretch intermetatarsal ligaments, which rebound elastically to draw the distal portions of the outer metatarsals together. This resulting distal unification does not have a counterpart in the horse carpus, although a long snap ligament absorbs shock to the entire forefoot.
Tensile keystone dynamics may explain the benefit of retention of multiple elements in the tyrannosaurid arctometatarsus, which contrasts with fused metapodia in ratites and in horses, bovids, cervids, camelids, giraffids, and other ungulates. A system of three bones and elastic ligaments may have imparted resilience and enhanced collective strength, properties diminished in a single metapodial element. The retention of multiple metatarsals as a stay against torsion may be paralleled in the Patagonian cavy, an agile cursorial rodent whose mesaxonic metapodia subtend an arch (pers. obs.). However, the metatarsals of the cavy lack the extremity of plantar angulation seen in the arctometatarsus, so the analogy is superficial and remains to be tested biomechanically.
A dynamically robust metatarsus is perhaps selectively logical in tyrannosaurids, which are much larger than most classically cursorial (fast-running) ratites and ungulates. Giraffes are potentially problematic to this view, because they are closer in mass to tyrannosaurids and have fused metapodia. As quadrupeds, giraffes have the advantage of lower loadings on the metapodia when trotting because two limbs share the load, although forces on each metapodium when galloping would be higher because the duty factor is low ( Alexander et al., 1977). Giraffes are also not as fast as might be expected from the extreme elongation of their limbs. The energy-absorbing metapodium of adult tyrannosaurids conceivably enabled them to outmatch giraffes in certain maneuvers or in linear speed, but such transtemporal comparisons are unproductively speculative.
The preceding discussion derives from an adaptationist perspective ( Gould and Lewontin, 1979). In contrast, phylogenetic and developmental contingency, rather than adaptation (Gould and Vrba 1982), also helps to explain the persistence of separate elements in the tyrannosaurid metatarsus. Giraffids, including the modern giraffe and okapi, inherited their metatarsal morphology from less specialized artiodactyls. Ratite birds inherited fused metatarsals from their avian ancestors and selective pressures for cursoriality need not be invoked to explain their ankylosed morphology. With this caveat in mind, we now explore arctometatarsus function in the context of performance and phylogeny.
Comparative Phylogenetic and Functional Implications
The tensile keystone model differs from kinematics likely evident in the foot of Allosaurus or other theropods with three largely autonomous metatarsals. As with humans ( Kerr et al., 1987), footfall loadings would cause their outer metatarsals to splay beyond their resting orientation, essentially spreading the foot apart. During deviations from linear locomotion, metatarsals would experience increased bending loads individually, rather than as part of a single structure, as predicted for the arctometatarsus ( Fig. 6 View Fig. 6 ). Broad-footed theropods are not uniform in metatarsus morphology ( Snively, 2000). None of these animals, however, display plantar constriction of MT III consistent with distal unification of the metatarsals, which would occur in the arctometatarsalian pes under the tensile keystone model.
The probable multiple origin of the arctometatarsus ( Sereno, 1999; Holtz, 2000) suggests it was not a legacy morphology, which was simply retained with no contemporary utility. Instead, it may have conferred a selective or performance benefit. Developmental and immediate functional advantages are not mutually exclusive. The correlation between a constricted third metatarsal and proportionally long metatarsus ( Holtz, 1995) suggests a developmental correspondence. Unfortunately, developmental hypotheses of this type are tenuously ad hoc. Perhaps the ontogenetic program for lengthened separate metatarsals reciprocally invoked proximal and plantar constriction of MT III in coelurosaurs, but tensile keystone dynamics evince more for the tyrannosaurid arctometatarsus than simply a developmental contribution to the lengthened foot.
Another possibility is that the tensile keystone morphology conferred heightened agility for a given body mass. As such, the arctometatarsus may have been broadly analogous to the stiffened tails of dromaeosaurid coelurosaurs ( Ostrom, 1969), which have been suggested as dynamic stabilizers. Because there was no anteriorly propulsive component to the elastic rebound of ligaments, third metatarsal constriction did not directly avail increased speeds. Instead, the unifying and shear-resisting properties of the arctometatarsus may have absorbed forces involved in linear deceleration, lateral acceleration, and torsion more effectively than the feet of other theropods. These forces are limiting factors to combat performance in humans (Snively: pers. obs. in open hand and weapons sparring) and the arctometatarsus may have imparted momentarily excessive construction ( Gans, 1974) for selectively crucial behaviors, such as predation or escape.
However, while the potential may have been present the employment and utility of increased agility in tyrannosaurids are no more directly testable than ontogenetic hypotheses. As with the evolutionary correlation between cursoriality and pursuit predation in theropods ( Carrano, 1998), alternate hypotheses must be explored. In addition, the tensile keystone model cannot be taken to indicate that tyrannosaurids behaved more dynamically than Allosaurus . Whether tyrannosaurids used the potential for higher maneuverability during prey capture, and how close these animals operated to safety limits, are untestable by observation. Conse- quently, definitive statements about comparative agility in theropods are premature. However, the tensile keystone model demonstrates, in one aspect of hindlimb function, potential benefits to agility in large arctometatarsalians.
CONCLUSION
Although the sample size of large, rare fossil organisms is notoriously small ( Kemp, 1999), the morphological evidence outlined above suggests significant dynamic differences between the metatarsi of tyrannosaurids and other large theropods. The tensile keystone model proposes that orientation and extent of ligaments in the arctometatarsus increased resistance to dissociation over that of other theropods, and yet allowed resiliency otherwise diminished in metapodia reduced to a single element, as in extant ratites, horses, or giraffids. These subhypotheses must be tested thoroughly in order to falsify or augment the overall model. Promising methods for further elucidating tyrannosaurid arctometatarsus function include dynamic computer modeling, quasi-static modeling ( Hutchinson, 2000), and finite element analysis of strain in the tyrannosaurid foot ( Snively and Russell, 2002).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |