
Testudo oughlamensis, Gmira & Broin & Geraads & Lefèvre & Mohib & Raynal, 2013
|
publication ID |
https://doi.org/10.5252/g2013n3a9 |
|
publication LSID |
lsid:zoobank.org:pub:1FFDA751-EDFE-40E6-8AAB-50AFAB78B78C |
|
persistent identifier |
https://treatment.plazi.org/id/38055C09-FB66-FF99-FC9F-F572FE36FD16 |
|
treatment provided by |
Marcus |
|
scientific name |
Testudo oughlamensis |
| status |
sp. nov. |
Testudo oughlamensis n. sp.
( Figs 2-6)
Testudo – Bailón 2000: 542. Testudo aff. kenitrensis – Lapparent de Broin 2000a: 62.
HOLOTYPE. — Collection INSAP. AaO-272, carapace incomplète: bord antérieur légèrement érodé aux périphériques 1 et 2 droites et à la nucale. Erosion des neurales 1 et 2, des parties arrière de la pleurale 2 et antérieure de la pleurale 3 droites, de la partie latérale des pleurales 2 et 3 gauches, et de l’arrière: majeure partie des pleurales 8, les suprapygales, la pygale et une grande partie de la bordure périphérique arrière (de la partie postérieure de la périphérique 7 gauche jusqu’à la partie postérieure de la périphérique 6 droite) et périphériques 2 à 5 gauches. Plastron sans les xiphiplastrons. Le moule interne est conservé par places permettant de suivre les contours de plaques manquantes.
PARATYPES. — Collection INSAP. Au moins dix individus. AaO-948: carapace incomplète et fragmentée représentée par la moitié gauche de la nucale avec la pleurale 1 gauche, une partie de la pleurale 1 droite, les neurales 3 quadrangulaire et 4 octogonale avec la partie proximale des pleurales 3 et 4 gauches et 3, 4 et 5 (partielle) droites, les neurales 7 et 8; attachées au plastron, les périphériques 3, 4 et 6 à 8 gauches et 3 et 6 à 8 droites avec la partie distale de la pleurale 8 droite; des fragments de pleurales paires et impaires; le plastron incomplet médialement (morsure animale probable par comparaison avec des blessures identifiées comme telles sur des spécimens actuels; coll. MNHN.F, RA, ZA) et sans le xiphiplastron droit. AaO-949: une carapace incomplète assez jeune et fragmentée représentée par la nucale avec les périphériques 1 et 2 gauches et droites et une partie de la pleurale 1 gauche, sept périphériques, trois fragments de pleurales (paires et impaires), la pygale, le plastron dont manque la région latérale de l’hypoplastron gauche et celle médiale de l’hypoplastron gauche et avec les périphériques 6 à 8 gauches et 5 et 6 droites; deux fragments de diaphyses d’un humérus et d’un coracoïde. Fragments isolés trouvés avec AaO-948 et AaO-949: deux neurales paire (6 e?) et impaire (7 e?); une périphérique 11 avec pleurales 8 gauche et 6 et 7 droites; deux périphériques du pont et deux périphériques postérieures; deux portions proximales et trois portions médianes de pleurales paires (2 e, 4 e ou 6 e) et un fragment distal de pleurale paire; deux portions distales de pleurales paire et impaire associées et un fragment distal de pleurale paire; trois fragments de pleurales impaires; un fragment distal de pleurale 8 gauche avec un fragment de plastron. AaO-293, périphérique du pont (5 e ou 6 e), périphérique postérieure (10e) et neurale 8; AaO-645, hypoplastron gauche; AaO-659, périphérique du pont (8 e gauche); AaO-672, périphérique du pont, 8 e gauche; pleurale 8 gauche (partie distale), épiplastron gauche et hypoplastron droit; AaO-673, -674, -675, trois épiplastrons avec leur entoplastron; AaO-676 et -677, deux fragments d’entoplastrons; AaO-678, cinq fragments de périphériques; AaO-679, fragment d’hypoplastron; AaO-680, fragment d’hypoplastron gauche; AaO-681, AaO-682, fragments d’hypoplastron gauche; AaO-683, hypoplastron gauche; AaO-684, fragment d’hypoplastron droit; AaO-685, fragment d’hypoplastron gauche; AaO-686, fragment latéral d’hyoplastron droit; AaO-687, fragment de nucale; AaO-688, xiphiplastron gauche; AaO-689, pygale; AaO- 690, périphérique 11gauche; AaO-691, périphérique 1 gauche; AaO-692, périphérique 11 gauche; AaO-693, entoplastron; AaO-694, pygale; AaO-695, périphérique 2 gauche; AaO-696, humérus droit partiel; AaO-697, périphérique 11 gauche avec pygale; AaO-698, pygale; AaO-699, pygale; AaO-2113, périphérique 1 gauche; AaO-2114, périphérique 9 gauche; AaO-2115, neurale 5; AaO-2116, périphérique 2 gauche; AaO-2117, fragment de pleurale 1 gauche; AaO-2118, fragment de pleurale 1 droite; AaO-2119 fragment de pleurale; AaO-2120 pleurale 8; AaO-2121, AaO-2122, périphériques du pont (5e-6e) gauche; AaO-2123, 16 lots de plaques comportant en tout 34 périphériques, 31 pleurales, 7 neurales et 17 fragments non situés; AaO-2124, trois périphériques antérieure, du pont et postérieure, jeunes; AaO-2125, trois périphériques postérieures jeunes; AaO-2127, épiplastron gauche; AaO-2129, périphérique 9 gauche; AaO-2130, périphérique 10 gauche; AaO-4788, nucale; AaO-4789, hyoplastron gauche; AaO-4790, hyoplastron droit. Des fragments associés dans un lot peuvent appartenir à un même individu.
ÉTYMOLOGIE. — En référence au gisement Ahl al Oughlam.
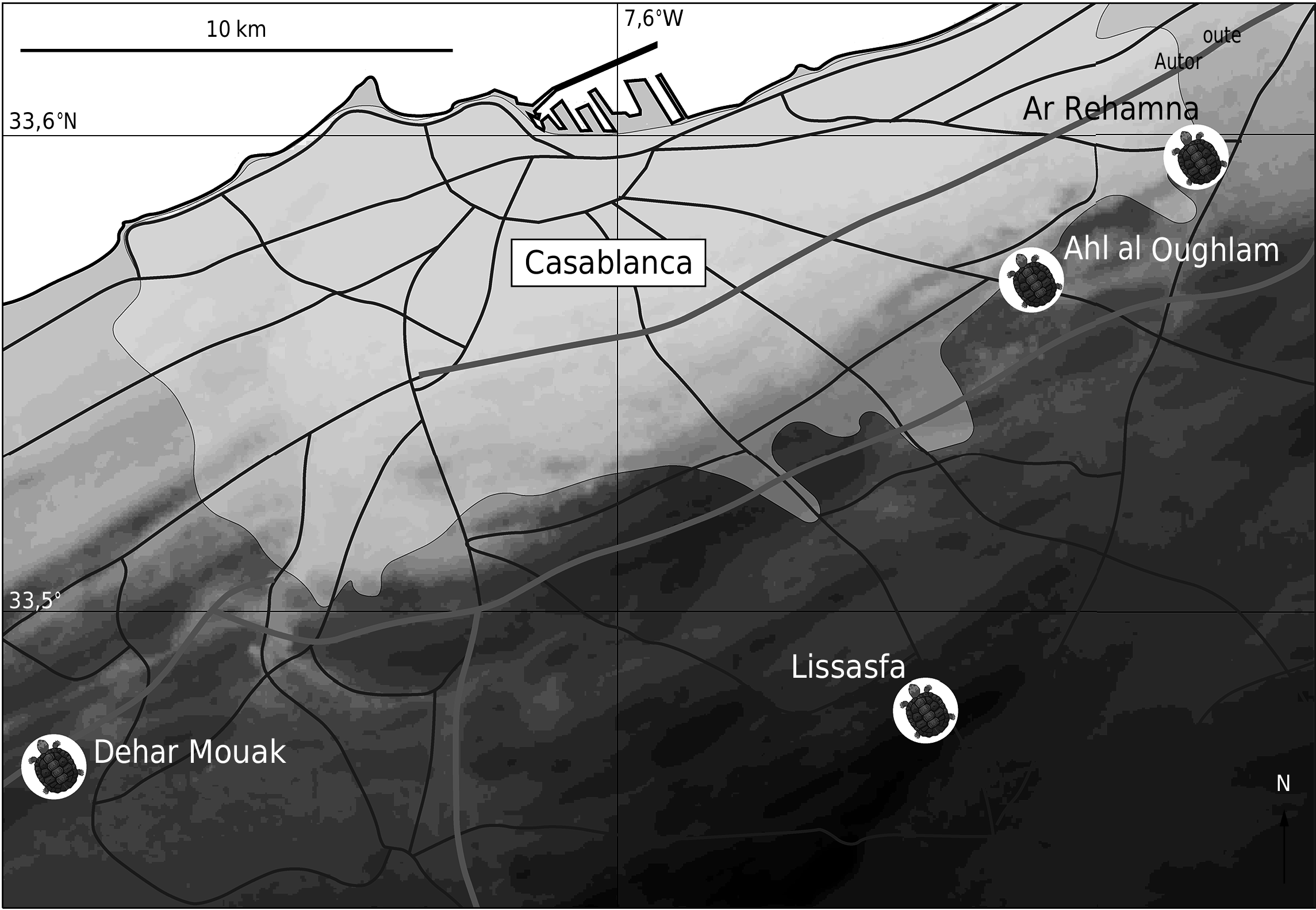
LOCALITÉ ET HORIZON STRATIGRAPHIQUE TYPES. — Remplissage continental dans la Carrière d’Ahl al Oughlam, située au sud-est de Casablanca, à gauche de la route S 106 allant de Casablanca à Tit Mellil. 33°34’11’’N, 07°30’44’’W. Cordons dunaires de la côte de Casablanca à Kenitra, Maroc. Unité morpho-sédimentaire de Sidi Messaoud. Carte géologique, feuille de Casablanca-Mohammedia ( Destombes 1987). Pliocène supérieur, âge estimé à 2,5 Ma ( Raynal et al. 1990; Geraads et al. 2010).
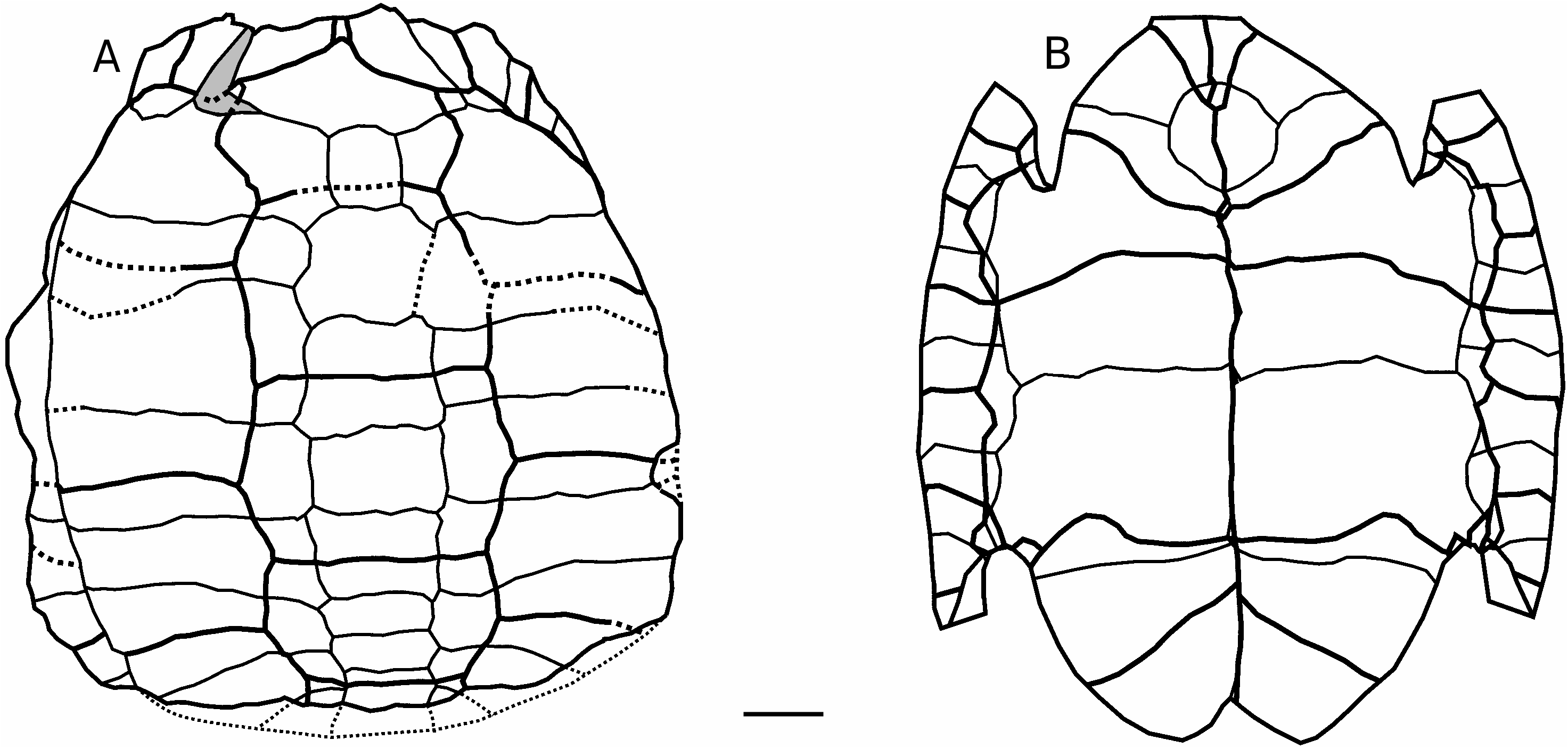
che; J, AaO-690, périphérique 11 gauche, face dorsale; K, AaO-699, pygale mâle, faces dorsale et latérale gauche. Abréviations: en, entoplastron; ep, bourrelet épiplastral; h, hyoplastron. La flèche indique l’encoche nucale à la périphérique 1. Échelle: 1 cm.
DIAGNOSE. — Espèce de Testudo formant notamment avec T. kenitrensis un groupe primitif par la bordure périphérique non redressée et la participation préservée de l’hypoplastron au lobe postérieur, à sillon abdominofémoral sinueux plus ou moins séparé de la charnière hypo-xiphiplastrale médialement et la rejoignant ou non latéralement et seulement sur le bord dorsal; dérivé par la petite taille de la carapace (ici 13 cm maximum) et le contour quadrangulaire dilaté médialement en vue dorsale, très arrondi en profil latéral: fortement convexe d’avant en arrière entre, d’une part, l’avant sous la nucale et les écailles vertébrales 1 et 2, et, d’autre part, l’arrière plus abruptement incliné, sous les écailles vertébrales 3 à 5, sans long plateau intermédiaire; par l’encoche nucale profonde, trapézoïdale, entre les périphériques 1 à la jonction des marginales 1 et 2; et par les vertébrales étroites. Formant avec T. kenitrensis un clade par le bourrelet épiplastral dorsal légèrement concave, épais, long et soulevé en corniche à l’arrière au dessus d’une petite poche gulaire et les anales longues, rapprochées médialement de la suture hypo-xiphiplastrale. Se distinguant de T. kenitrensis par le contour plus dilaté médialement en vue dorsale et les écailles vertébrales 2, 3 et 4 relativement plus étroites; le bord antérieur de la nucale en saillie vers l’avant au milieu de l’encoche; la vertébrale 1 recouvrant ou non les coins de la nucale, les bords latéraux de cette vertébrale 1 variant de rectilignes (divergents vers l’avant) à légèrement arrondis et à sinueux (vertébrale en lyre), alors que la vertébrale en lyre est constante chez T. kenitrensis ; le plastron à lobe antérieur plus allongé aux épiplastrons et le bourrelet épiplastral dorsal un peu moins long; la relativement plus étroite petite saillie gulaire par rapport aux humérales; le sillon fémoro-anal plus rapproché médialement de la suture hypo-xiphiplastrale et la présence de deux écailles inguinales au lieu d’une.
DESCRIPTION
Holotype
Longueur préservée: 106 mm (estimée à 120 mm maximum); largeur: 101 mm; hauteur: 65 mm. Vertébrale 3, largeur: 4 mm sur longueur: 28 mm. Bien qu’incomplet, c’est le seul spécimen montrant la forme globale de la carapace, apomorphe ( Figs 2; 3 View FIG ). Sa partie postérieure manquant et sa partie antérieure étant légèrement érodée, sa description est complétée par les paratypes.
Carapace dorsale. Le spécimen est un peu déformé et déjeté sur la droite, la bordure latérale apparaissant de ce fait un peu à gauche vers l’arrière ( Figs 2; 3 View FIG ), et le plastron est un peu enfoncé sous la dossière mais ses éléments sont plats, pouvant indiquer qu’il s’agit d’une femelle. L’épaisseur des plaques et des annuli de croissance montre qu’il s’agit d’un spécimen bien adulte. Le profil de la carapace en vue latérale ( Fig. 2A 4 View FIG ) montre une forme courte avec un contour arrondi, présentant une pente forte vers l’avant et une un peu plus forte à l’arrière, avec un court plateau intermédiaire à peu près horizontal sous la vertébrale 3, avec juste deux petites bosselures aux neurales 2 et 4 (sous les vertébrales 2 et 3) et de petites bosses aussi sous la vertébrale 4 et médialement sous les costales 2 et 3. La face dorsale montre également un contour arrondi, de chaque côté. En vue dorsale, la bordure périphérique antérieure (la postérieure non conservée) suit la pente du disque pleural sans se relever vers son extrémité. La nucale est un peu incomplète à gauche et séparée postmortem de la périphérique 1 gauche; elle est plus large que longue, hexagonale resserrée à l’arrière au contact avec la neurale 1 (la plus étroite de la série neurale). Le bord antérieur de la dossière forme une encoche entre les périphériques 1 (aux sillons entre marginales 1 et 2) avec la nucale en retrait ( Fig. 2A2), encoche amoindrie ici par l’érosion du bord des plaques, mais complète sur le spécimen AaO-249 ( Fig. 4B View FIG ) et estimable d’après la nucale et les périphériques 1 et 2 isolées ( Fig. 2 B-D). Les périphériques antérieures l et 2 sont longues, étroites dans leur extrémité postérieure (médiale) et s’élargissent vers l’avant. Leur bord libre externe, complété par les paratypes ( Figs 2; 4 View FIG ), était saillant en pointe à la jonction des écailles marginales 1-2 et 2-3 respectivement. Les périphériques antérieures sont en pente oblique suivant la pente du disque pleural, sans se redresser vers leur extrémité; les périphériques du pont étaient verticales: comme le spécimen a été déformé post-mortem et fléchi sur la gauche, les périphériques du pont apparaissent artificiellement en partie à gauche. Les neurales 2 à 5 sont différenciées mais très irrégulières: neurale 1 (manquante mais estimée par sa place entre les plaques environnantes) quadrangulaire; neurale 2 octogonale (son bord droit manquant); 3 e heptagonale (le petit côté latéro-antérieur gauche manquant); 4 e pentagonale avec un petit côté latéro-postérieur droit; 5 e hexagonale avec un petit côté latéro-antérieur gauche et un petit côté latéro-postérieur droit; 6 e pentagonale avec un petit côté latéro-antérieur gauche; 7 e hexagonale avec petits côtés latéro-anté- rieurs; 8 e hexagonale avec petit côté latéro-postérieur droit mais petit côté antérieur gauche aligné sur le petit côté postérieur gauche (séparés par la jonction pleurales 7 et 8). La différentiation irrégulière des neurales entraîne une irrégularité des pleurales dans leur longueur respective. Les pleurales 2 à 6 sont alternativement plus courtes et plus longues (d’avant en arrière) médialement et plus longues et plus courtes latéralement, mais irrégulièrement à cause de l’irrégularité des neurales, et les pleurales 2 droite et 3 gauche sont particulièrement longues. Les périphériques 3 à 7 du pont, partiellement érodées (surtout à gauche), sont hautes, leurs parties dorsale et ventrale étant très ouvertes en un angle de plus de 90° ( Fig. 2A 4 View FIG ); leur base ventrale est un peu enfoncée avec le plastron.
Écailles: la cervicale est assez longue sur la nucale (23 %); très étroite dorsalement, elle s’étrécit un peu à l’avant et son bord arrière est concave; elle est un peu plus large ventralement. La 1 re marginale est un peu plus courte que la périphérique 1; les marginales suivantes sont aussi longues que les périphériques, la suture périphériques-pleurales coïncidant avec le sillon marginales-costales. Les quatre écailles préservées de la série vertébrale sont plus larges que longues, hexagonales, mais la série est relativement modérément large; la largeur sur longueur de la vertébrale 3 (la plus large) est de 142 % (152-155 % chez T. graeca ). La 1 re est partiellement endommagée médialement; elle est en forme de lyre (bords latéraux sinueux): à partir de l’arrière, ses bords latéraux divergent vers l’avant, s’arrondissent en se rapprochant (étrécissement de l’écaille) pour continuer en s’écartant à peine et arriver sur la nucale en ne recouvrant pas les coins postéro-latéraux. Le rebord de la partie visible ventralement des marginales antérieures et de la cervicale est long ( 14 mm à la cervicale). Une écaille axillaire est visible à droite dans sa partie située sur la périphérique 3 (complète sur le spécimen AaO-948, Fig. 4A View FIG 2). Les annuli de croissance des écailles sont particulièrement visibles sur les pourtours pleural et périphérique droits.
Plastron. Longueur préservée (de l’avant des épiplastrons à l’arrière des hypoplastrons): 93 mm; longueur et largeur du lobe antérieur: 56 et 37 mm; largeur et longueur de la saillie gulaire: 22 mm et 5 mm; longueur et hauteur du bourrelet épiplastral dorsal: 22 mm et 16 mm; largeur des deux gulaires à l’arrière du bourrelet épiplastral dorsal: 28 mm. Longueur du pont: 64 mm. Le lobe antérieur est trapézoïdal avec une rupture de pente: ses bords latéraux sont d’abord un peu convergents (angle à l’encoche axillaire assez fermé, c. 35°) puis ses bords convergent davantage pour se resserrer encore à la saillie gulaire, nette. Il est nettement plus étroit que le lobe postérieur.Le lobe antérieur est relativement long du fait des épiplastrons allongés: le rapport de la longueur du bord de l’épiplastron à la longueur du bord latéral du lobe antérieur est de 69 % ( T. graeca : 48 %). Dorsalement, les épiplastrons s’élèvent et se courbent vers l’arrière un peu en corniche jusqu’au-dessus de l’avant de l’entoplastron: ce bourrelet est élevé et s’incurve un peu, se faisant légèrement concave. L’entoplastron a un aspect rhombique, côtés latéro-postérieurs arrondis, aussi long que large; il est petit dans le lobe antérieur et atteint ou dépasse à peine postérieurement le fond des encoches axillaires. Le pont est long et massif. Les hypoplastrons participent un peu au lobe postérieur. Les xiphiplastrons manquent mais le bord postérieur libre des hypoplastrons a une surface denticulée qui montre l’existence d’une charnière hypo-xiphiplastrale.
Écailles: les gulaires sont saillantes latéralement par rapport aux humérales mais elles ne sont pas en relief ventralement par rapport à ces dernières; ventralement triangulaires, elles s’étrécissent un peu en coin, en s’incurvant, atteignant la pointe de l’entoplastron. Dorsalement, elles couvrent le bourrelet épiplastral dorsal en s’élargissant un peu vers l’arrière. Le sillon huméro-pectoral contourne l’entoplastron, éloigné latéralement et plus près médialement tout en restant bien à l’écart de la plaque. Médialement derrière l’entoplastron, se font de plus en plus grandes, se multipliant par deux en longueur chaque fois: les humérales, les pectorales et les abdominales. Les abdominales ne parviennent pas à la suture hypoplastrale postérieure médialement et forment une sinuosité convexe de chaque côté. L’abdominale droite est incomplète latéralement mais la gauche est préservée: elle rejoint presque la suture-charnière latéralement en arrière d’une petite écaille inguinale hypoplastrale. Des annuli de croissance des écailles sont visibles sur l’hypoplastron.
Paratypes
Ils sont indispensables pour compléter l’holotype et pour appréhender la variabilité spécifique. Les caractères essentiels de l’holotype sont confirmés par les plaques isolées et les spécimens AaO-948 et AaO-949 ( Figs 2; 3B View FIG ; 4-5 View FIG ). Les plastrons de ces deux spécimens sont plats et devaient appartenir à des femelles, ce que confirme la pygale conservée du second, non épaissie et peu recourbée.
Carapace dorsale. La nucale complète est préservée par les spécimens AaO-4788 ( Fig. 2D), AaO-948 ( Fig. 4A4 View FIG ) et le plus jeune AaO-949 ( Fig. 4B View FIG 1 View FIG , B 2), celui-ci montrant l’encoche nucale dans son intégralité. Les plaques neurales isolées et celles de AaO-948 montrent comme l’holotype la différenciation de neurales médianes, régulière dans la partie préservée chez AaO-948: 3 e quadrangulaire, 4e octogonale et 5e quadrangulaire, alors que cette partie est irrégulièrement différenciée chez l’holotype. Plusieurs périphériques antérieures sont préservées dont la première d’AaO-691 ( Fig. 2C) et d’AaO-949 ( Fig.4B View FIG ) avec la seconde d’AaO-695 ( Fig. 2B) et d’AaO-649 ( Fig. 4B View FIG ): elles montrent aussi l’encoche profonde trapézoïdale du bord antérieur, avec les périphériques 1 saillantes à bord médial encoché (concave), et la nucale projetée vers l’avant au milieu; les plaques périphériques 1 et 2 et la nucale forment une pointe à la jonction externe des marginales entre elles et avec la cervicale. La périphérique 2 s’épaissit postérieurement au contact de la 3 e qui ouvre le pont. Le pont est bien visible chez AaO-948, AaO-949 et plusieurs plaques isolées: les plaques du pont sont hautes à angle obtus au milieu du pont. La périphérique 3 porte le contrefort axillaire de l’hyoplastron dressé verticalement et un peu en oblique contre sa moitié antérieure épaissie et le début du pont à sa moitié postérieure, s’ouvrant largement sur les périphériques 4 à 6 ouvertes en angle obtus. La périphérique 7 ferme le pont et porte dans sa moitié postérieure le contrefort inguinal hypoplastral dressé verticalement et débordant un peu sur le bord ventroantérieur de la périphérique 8, verticale, dont la face externe est un peu bombée et dont le bord ventral est libre comme les suivantes. Les postérieures sont conservées en plusieurs exemplaires: la périphérique 9 ( Fig. 5O 1 View FIG ), également bombée extérieurement, se fait plus courte et s’épaissit postérieurement comme l’avant de la 10 e ( Fig. 5O 2) laquelle s’amincit postérieurement à l’épaisseur de la 11 e et de la pygale. Ces plaques postérieures sont verticales et ne sont pas redressées extérieurement à leur extrémité libre. Plusieurs plaques isolées appartiennent à des jeunes: elles sont relativement plus courtes et plus minces ( Fig. 5N) comme chez AaO-949. De la partie postérieure, il manque la ou les suprapygales.D’après les pleurales 8 préservées et les pygales, la ou les suprapygales formaient un bloc trapézoïdal à bord postérieur transversal. La pygale est conservée en plusieurs exemplaires ainsi que les périphériques postérieures, du pont à la 11 e ( Figs 2; 4 View FIG ). La pygale est trapézoïdale à grand côté antérieur (qui contactait la suprapygale) et était encadrée par les périphériques 11 formant un coin aigu médiopostérieurement contre elle; la pygale varie un peu dans ses proportions ( Figs 2 E-K; 4B6): certaines pygales, attribuées aux mâles, sont plus courbées, plus larges et plus épaisses ( Fig. 2I, K) que les autres ( Fig. 2H). Les pygales sont allongées par rapport à la condition primitive de la famille: leur longueur atteignait celle des périphériques ( Fig. 2E, J, K), jusqu’au bord postérieur de la suprapygale prolongeant le bord des pleurales 8 sur une même ligne; de sorte que, comme antérieurement entre plaques et sillons à partir de la 2 e périphérique, la suture périphériques/pleurales-suprapygale coïncidait avec le sillon marginales/costales-vertébrale 5.
Écailles: les paratypes montrent la variabilité de l’écaille vertébrale 1: ses bords latéraux sont en lyre sur l’holotype, divergents puis presque parallèles à l’avant chez AaO-948 et à peine arrondis vers l’extrémité antérieure chez AaO-949. Le spécimen de la nucale isolée AaO-4788 pouvait aussi avoir eu une vertébrale en lyre. Les bords de cette vertébrale ne couvrent pas les coins de la nucale chez l’holotype AaO-272, et à peine chez AaO-949 et les nucales isolées AaO-4788 et AaO-687, mais la vertébrale 1 les couvre complètement chez AaO- 948. Les pygales ont une surface lisse, sans sillon médian ce qui indique la présence d’une supracaudale simple (marginales 12 fusionnées).
Plastron. Comme sur l’holotype, le lobe antérieur est plus étroit que le postérieur avec des angles axillaires relativement fermés; les épiplastrons sont tous allongés, à saillie gulaire nette, le bourrelet épiplastral dorsal est toujours légèrement concave. Un seul spécimen sur huit montre que le bourrelet épiplastral ne couvre pas toujours l’entoplastron ( Fig. 5D). Les hypoplastrons forment tous charnière avec les xiphiplastrons comme chez l’holotype, qu’ils soient femelles (par ex. AaO-948 et AaO-949) ou mâles (par ex. AaO-672; Fig. 5L). Les hyoplastrons sont épaissis médialement aux processus axillaires et les hypoplastrons et xiphiplastrons sont épaissis latéralement au maximum dans leur zone de contact, à la limite médiale du sillon des écailles fémorales. Tous les hypoplastrons participent un peu au lobe postérieur en avant des xiphiplastrons. Le lobe postérieur est large et ses bords sont arrondis jusqu’à la pointe xiphiplastrale. Les xiphiplastrons sont aussi larges que longs ou un peu plus larges ( Fig. 5M), resserrés un peu ou non au sillon fémoro-anal, mais devenant plus convergents vers la pointe. Ils sont aussi épaissis au maximum à l’avant de leur pointe xiphiplastrale, au sillon médial des écailles anales. L’encoche anale est triangulaire, plus large que longue mais relativement profonde ( Figs 4A View FIG 1 View FIG , A 2, B 3 View FIG , B 4 View FIG ; 5M); seuls trois spécimens sont connus par leur xiphiplastron dont les deux présumés femelles (AaO-948 et AaO-949) et le troisième (AaO-688, Fig. 5M) n’est pas affirmé comme mâle ou femelle, ne permettant pas de déceler un éventuel dimorphisme sexuel.
Écailles: les gulaires atteignent juste l’entoplastron ou le recouvrent un peu dorsalement. Elles s’élargissent à peine vers l’arrière sur le bourrelet épiplastral dorsal comme chez l’holotype. Il y a une écaille axillaire triangulaire, allongée à cheval sur la périphérique 3 et l’hyoplastron, visible sur AaO-948 ventralement. Le sillon huméro-pectoral contourne toujours l’entoplastron sans s’en approcher, avec une légère sinuosité convexe latérale et médialement en arrondi ou plus directement. La sinuosité plus ou moins latérale ou médiale du sillon pectoro-abdominal sur l’hyoplastron est faible. La sinuosité du sillon abdomino-fémoral est variable, de faible (sillon presque parallèle à la charnière hypo-xiphiplastrale) à forte, le sillon s’approchant le plus fortement de la suture médialement dans un seul cas sur 12 ( Fig. 5L 1 View FIG ) et restant séparé de la suture latéralement en vue ventrale; il rejoint ou non la suture dorso-latéralement (non visible ventralement). Les anales s’étendent médialement vers l’avant vers la suture hypo-xiphiplastrale, le sillon fémoro-anal restant à peu près parallèle au bord de l’encoche anale, laquelle est relativement profonde. La longueur des anales sur le xiphiplastron est plus grande que la longueur des abdominales sur l’avant des xiphiplastrons comme aussi sur l’arrière de l’hypoplastron. Il y a deux écailles inguinales, visibles ventralement sur certains spécimens dont AaO-948, une petite hypoplastrale et une plus grande plus facilement visible, à cheval sur la périphérique 7 et l’hypoplastron, ventralement; elles sont séparées ventralement, se joignant dorsalement.
Humérus, AaO-696 ( Fig. 6A View FIG ). Il est incomplet distalement et aux trochanters. La diaphyse est recourbée, les trochanters sont écartés par une fosse intertrochantérique en U plus large que chaque trochanter. La tête est arrondie.
COMPARAISON
La détermination de la nouvelle espèce de Testudo d’Ahl al Oughlam et des spécimens décrits plus loin est basée sur leur comparaison avec les espèces actuelles de Testudo , les spécimens du Pléistocène- Holocène du Maroc déjà décrits ( Gmira 1995) dont l’espèce T. kenitrensis Gmira, 1993 , du Pléistocène moyen de Kénitra (au nord de Rabat) et les autres restes de Testudo fossiles du Maghreb ( Lapparent de Broin 2000a). Les tortues fossiles du Maroc attribuées à l’espèce actuelle T. graeca (s.s.) comportent les spécimens décrits in Gmira (1995) en tant que Testudo graeca graeca . Depuis l’étude de Gmira (1995) de nouveaux travaux modifient les rapports entre les espèces du genre et les rapports entre les genres de la sous-famille. Les études cladistiques menées à ce jour ( Gmira 1995; Lapparent de Broin et al. 2006b, c) ne sont pas concluantes pour les relations phylétiques précises des genres distingués au sein d’un Testudo s.l. paléarctique subdivisé, du fait de l’insuffisance de caractères fournis pas les carapaces osseuses seules et de la grande quantité d’homoplasies possibles connues. Elles ont cependant permis de mieux apprécier les différences entre les genres correspondant aux subdivisions reconnues. Conformément à ces études, le genre Testudo est limité ici aux petites espèces paléarctiques à charnière hypo-xiphiplastrale chez les deux sexes et à bourrelet épiplastral élevé. Sont formellement exclues du genre Testudo les espèces sans charnière dans les deux sexes appartenant: 1) au genre paléarctique oriental Agrionemys Khozatsky & Mlynarski, 1966 ; 2) au genre fossile Paleotestudo Lapparent de Broin, 2000 , au sens large, en incluant le groupe promarginata ( Broin, 1977) outre l’espèce type canetotiana Lartet, 1851 et P. mellingi ( Peters, 1869) (voir Lartet 1851; Peters 1869); et 3) au genre « Eurotestudo » in Lapparent de Broin et al. 2006a-c (espèces fossiles et actuelles) i.e. Chersine Merrem, 1820 , espèce type Testudo hermanni Gmelin, 1789 , not Chersina Gray, 1831 ( Gray 1831a, b) espèce type Testudo angulata Schweigger, 1812 . Il a été montré que ( Bour & Ohler 2008), n’étant pas citée dans la publication originale du genre Medaestia Wussow, 1916 , l’espèce Testudo hermanni ne pouvait être attribuée à ce dernier genre (contrairement à l’une des propositions de Fritz & Kraus 2008) mais seulement à Chersine . Il a été prouvé que les lignées Chersine (« Eurotestudo » in Lapparent de Broin et al. 2006a-c), Agrionemys et Testudo sont distinctes depuis le Miocène supérieur, Vallésien, MN 10, au moins ( Lapparent de Broin et al. 2006b, c; Garcia et al. 2011). Au sein du genre Testudo s.l., les divisions et regroupements en espèces ou sous-espèces actuelles sont basées souvent sur des caractères dont beaucoup proviennent de la taille et de la morphologie générale, mais surtout des écailles et des couleurs de la tête, de la carapace et des membres, de sorte que les caractères ne sont pas conservés chez les fossiles; bien de ces caractères sont souvent reconnaissables seulement par une étude statistique avec un échantillon important ( Perälä 2002a, b; Pieh & Perälä 2004; Lapparent de Broin et al. 2006b, c). Dès les premières études ( Gmira 1992, 1993a, b, 1995) a été montrée la nécessité d’avoir, en ce qui concerne les fossiles comme les actuels, un échantillon suffisant de squelettes, ce qui n’est pas souvent le cas. Le groupe T. graeca s.s. est considéré ici comme incluant les sous-espèces actuelles du Maghreb, s’étendant du Maroc à la Libye: Testudo g. graeca Linnaeus, 1758 ; T. g. cyrenaica Pieh & Perälä, 2002 ; T. g. soussensis Pieh, 2000, T. g. marokkensis Pieh & Perälä, 2004; T. g. lamberti Pieh & Perälä, 2004; T. g. nabeulensis Highfield, 1990. Les spécimens fossiles du Maroc attribués à Testudo graeca sont déterminés à l’aide des squelettes de spécimens actuels ( Algérie, Maroc) des collections du MNHN, dont ceux récoltés dans les régions de Fez et de Rabat; pour ceux-ci, la sous-espèce possible serait T. g. marokkensis mais les squelettes à notre disposition ne permettent pas vraiment d’attribution précise en regard des définitions des sous-espèces sur les caractères externes.
Chez T. oughlamensis n. sp. la plupart des caractères sont généralisés et communs à plusieurs genres de Testudininae incluant Testudo et représentant plusieurs lignées dans le monde ( Broin 1977; Lapparent de Broin 2000b, 2002, 2003, 2008; Lapparent de Broin et al. 2006b, c). Ces caractères généralisés sont soit présents à la base de la sous-famille, soit communs par synapomorphie ou par homoplasie chez certains genres à partir d’un certain grade évolutif, mais seule une étude cladistique complète des Testudinidae mettrait en évidence leur répartition: 1) la forme générale de la carapace (dossière et plastron, plaques et écailles); 2) la présence d’écailles axillaires et inguinales (primitive); 3) la forme générale de la plaque nucale; 4) la différentiation des neurales avec une première neurale quadrangulaire (primitivement elle est hexagonale à petits côtés postérieurs) précédant quelques plaques octogonales et carrées; la différenciation des neurales en plaques octogonales et carrées n’est pas stabilisée ici d’après l’holotype; 5) la forme du plastron avec les gulaires triangulaires longues (primitivement elles sont très courtes); 6) la morphologie des étroits et courts contreforts plastraux dressés le long des périphériques du pont; 7) le lobe postérieur plus large que l’antérieur; et 8) le pont long.
La présence de pointes aux extrémités des sillons des marginales est un caractère représenté chez certains Testudininae dans les lignées paléarctiques dès l’Éocène supérieur et l’Oligocène dont « l’ Ergilemys Čkhikvadze, 1972 » de France ( Broin 1977, 1978; Lapparent de Broin 2000b; Lapparent de Broin et al. 2006b, c) et certains petits endémiques africains actuels tels Homopus Duméril & Bibron, 1834 , et Kinixys Bell, 1827 , ainsi que Namibchersus namaquensis (Stromer, 1926) du Miocène inférieur ( Lapparent de Broin 2003, 2008), accompagnant éventuellement une encoche du bord antérieur. La coïncidence des sillons des écailles costales-vertébrale 5/marginales-supracaudale avec les sutures pleurales-suprapygales/périphériques-pygale sur le pourtour de la dossière est un caractère dérivé par rapport au stade du morphotype le plus primitif (celui de Manouria impressa (Günther, 1882)) et au stade du morphotype « Geochelone » ( Lapparent de Broin 2003, 2008): la pygale est élevée ici jusqu’au niveau des périphériques et sa suture proximale avec la suprapygale coïncide avec la limite postérieure de la vertébrale 5. Ce caractère est notamment partagé par Paleotestudo , Chersine (« Eurotestudo » in Lapparent de Broin et al. 2006a-c), Testudo et « Testudo » semenensis Bergounioux, 1955 du Miocène supérieur de Djebel Sémène ( Tableau 1) de Tunisie (voir Bergounioux 1955). Dans l’ancien monde, la configuration suprapygale – pygale de ces derniers est proche aussi de celle d’ Agrionemys (mais avec une différentiation dans le raccourcissement médian de la vertébrale 5 chez ce dernier) et de celle des petits endémiques africains (mais avec d’autres particularités des suprapygales et du sillon de la vertébrale 5) ( Gmira 1995; Lapparent de Broin 2003; Lapparent de Broin et al. 2006b, c). La fusion des marginales 12 en une supracaudale est apparue à partir de l’Éocène supérieur-Oligocène en Europe occidentale, comme chez l’oriental Agrionemys et aussi chez d’autres Testudininae ; une redivision progressive de la supracaudale se produit aux Pliocène-Quaternaire chez Chersine , soit d’un seul côté, soit des deux côtés (dorsal et ventral) comme le plus souvent chez les spécimens actuels. La pygale de T. oughlamensis n. sp. est recourbée vers l’intérieur comme chez la plupart des espèces de Testudininae néogènes et au moins chez les mâles actuels, sauf chez T. marginata où la bordure postérieure s’étale plus largement y compris au niveau de la pygale chez les mâles comme chez les femelles.
T. oughlamensis n. sp. et T. kenitrensis partagent avec la majorité des espèces de Testudininae (primitivement) la sinuosité latérale modérée du sillon pectoro-adominal.Toutefois T. marginata Schoepff, 1789 et T. kleinmanni Lortet, 1883 se distinguent des autres espèces de Testudo , y compris les fossiles du Maroc décrits ici, par l’infléchissement médian des pectorales beaucoup plus fort, celles-ci rejoignant presque la suture hyo-hypoplastrale; par ailleurs T. marginata et T. kleinmanni se distinguent l’une de l’autre par d’autres caractères absents chez les Testudininae du Maroc, dont la forte encoche nucale en V chez T.kleinmanni , et la longue carapace étroite et l’étalement de la bordure postérieure chez T. marginata ( Lapparent de Broin et al. 2006b, c).
La petite taille de l’entoplastron est un caractère dérivé commun à Paleotestudo , Testudo et Chersine , et parfois aussi à certains endémiques africains où il a tendance à se réduire encore plus ( Lapparent de Broin 2003, 2008).
Testudo oughlamensis n. sp. et T. kenitrensis forment un clade avec les autres espèces de Testudo , défini principalement par la présence d’une charnière hypo-xiphiplastrale chez tous les individus ( Lapparent de Broin et al. 2006b, c) avec de ce fait le raccourcissement des hypoplastrons à l’arrière, la proximité du sillon abdomino-fémoral et de la charnière médialement, avec la possibilité de leur contact tout à fait latéralement, dorsalement au moins. La présence d’une charnière hypo-xiphiplastrale existe aussi chez des spécimens isolés d’espèces de Testudininae indéterminées de l’Oligocène et de l’Aquitanien de France attribuées avec doute à « Ergilemys » sp., avec une conjonction différente de caractères des éléments isolés dont la pygale large et courte et les marginales 12 divisées (en partie au moins) primitivement d’une part, les xiphiplastrons atteignant les encoches inguinales (seule participation du xiphiplastron au lobe postérieur ( Broin 1977: pl. 25, figs 11, 12), ce qui est plutôt rare chez Testudo ) et le bourrelet épiplastral long et aplati apomorphiquement d’autre part ( Broin 1977, 1978; Lapparent de Broin et al. 2006b, c).
Testudo oughlamensis n. sp. et T. kenitrensis diffèrent des espèces de Testudo actuelles par deux caractères primitifs: 1) la bordure périphérique postérieure ne s’étale pas «en une jupette » qui serait visible dorsalement chez T. kenitrensis et, en l’absence de celles de l’holotype de T. oughlamensis n. sp., les périphériques postérieures isolées ( paratypes) bien verticales indiquent qu’il en allait de même dans cette dernière espèce. Il en est de même chez Paleotestudo . Une éventuelle différentiation sexuelle chez les espèces actuelles du Maghreb dans l’importance de l’étalement n’est pas évidente sur le squelette osseux: dans l’ensemble une fois les écailles enlevées, les spécimens adultes de T. graeca du Maroc examinés, et plus précisément ceux de Rabat à Fez, ont un bord postérieur osseux variable d’à peine redressé postérieurement aux périphériques 10 et 11 à nettement étalé aux périphériques 11 et redressé à la pygale chez la femelle (pygale bombée mais aussi visible dorsalement chez le mâle); cet étalement est visible dorsalement, par exemple chez les spécimens de T. graeca du Pléistocène supérieur du Maroc ( Gmira 1995: pl. VIIA) et chez la plupart des autres espèces actuelles de Testudo avec une grande variabilité suivant celles-ci: étalement postérieur beaucoup moins marqué chez un petit spécimen de Testudo cyrenaica de Libye femelle que chez un grand spécimen femelle de T. ibera de Turquie par exemple; un extrême étalement, très dérivé, se produit chez T. marginata mâle et femelle; 2) La participation primitive de l’hypoplastron au lobe postérieur est constante dans le clade kenitrensis , le sillon abdomino-fémoral, sinueux, restant séparé de la charnière sur la face ventrale, latéralement et médialement; cette participation tend à disparaître totalement chez les spécimens actuels. Chez de nombreux spécimens actuels de T. graeca du Maroc (dont REP 21, coll. en dépôt au MNHN.F, 17 cm de long) et d’autres d’espèces plus orientales ( T. ibera , T. marginata ), le sillon abdomino-fémoral contacte largement la suture médialement ( Lapparent de Broin 2002: fig. 19) et se confond souvent presque totalement avec la charnière; l’hypoplastron est raccourci jusqu’à ce qu’il ne participe pratiquement plus au lobe postérieur et que le xiphiplastron atteigne le fond de l’encoche inguinale. Pour ce dernier cas, il en est de même chez les éléments isolés attribués à « Ergilemys » ( Broin 1977: pl. 25, fig. 11; pl. 28, fig. 7) (voir ci-dessus).
La taille réduite est dérivée mais elle n’est pas unique aux espèces fossiles marocaines: T. oughlamensis n. sp. comme T. kenitrensis sont d’une petite taille, plus petite mais assez proche de celle de la population actuelle du NW et du centre du Maroc, T. g. marokkensis Pieh & Perälä, 2004. La taille est encore plus petite chez T. kleinmanni d’Égypte.
L’angle des encoches axillaires est variable chez les actuels Testudo , variant d’aussi fermé dans le clade kenitrensis que chez T. graeca à plus ouvert ( T. kleinmanni ), comme chez Mauremys le géoémydiné présent aussi à Ahl al Oughlam.
Testudo oughlamensis n. sp. et T. kenitrensis forment un clade kenitrensis qui diffère de toutes les autres espèces de Testudo par la présence du bourrelet épiplastral élevé et en corniche mais à surface dorsale légèrement concave. Chez les autres espèces de Testudo , ce bourrelet est aussi élevé mais à surface convexe comme chez d’autres Testudininae ( Lapparent de Broin et al. 2006b: fig. 3) et il est plus court par rapport à la largeur de la saillie gulaire chez les espèces actuelles. Ce caractère de la concavité du bourrelet élevé est primordial pour caractériser le clade. Il est considéré ici comme un caractère dérivé à partir du stade à bourrelet convexe de Testudo considéré comme plus primitif: toutefois celui-ci est lui-même dérivé pour les Testudinidae . Le bourrelet convexe est plus développé que dans les stades de base à bourrelet non élevé et largement concave tel celui préservé chez l’actuel Manouria impresssa (Günther, 1882) ( Lapparent de Broin et al. 2006b: fig. 3a) et encore proche de celui des Geoemydinae, et tel celui un peu plus dérivé du genre indéterminé à ce jour, représenté par « Achilemys » cassouleti Claude & Tong, 2004 et « Hadrianus » sp. in Broin (1977) de l’Éocène inférieur de France (voir Claude & Tong 2004), génériquement nommés de façon inappropriée. Ce stade est aussi différemment développé du long bourrelet aplati des « Ergilemys » de France ( Broin 1977: figs 109, 110; pl. 27: fig. 11; pl. 29). Le stade convexe de Testudo est partagé avec les genres occidentaux Chersine (« Eurotestudo » in Lapparent de Broin et al. 2006a-c) (Pliocène-Actuel) et Paleotestudo , avec l’oriental Agrionemys et indépendamment (avec des variations dans les proportions) avec d’autres genres de Testudininae comme par exemple en Afrique dans d’autres lignées telle celle de Stigmochelys Gray, 1873 ( Lapparent de Broin 2003, 2006b). À la différence de Testudo , tous ces genres sont tous toujours dépourvus de charnière plastrale (au moins chez les mâles car les femelles adultes de Chersine hermanni hermanni peuvent en avoir une, facilitant l’oviposition). Mais chez Chersine et Agrionemys l’extension du bourrelet ne va pas jusqu’au-dessus de l’entoplastron, la poche gulaire est antérieure à l’entoplastron et faible quand elle existe chez Chersine (dans les populations de l’espèce Chersine hermanni du Quaternaire de Lunel-Viel et de Soave ) et il y a toujours l’absence de charnière et la division secondaire de la supracaudale ainsi que d’autres particularités pour distinguer le groupe hermanni ; tandis que d’autres caractères dont la forme du lobe antérieur à grand entoplastron et la grande hauteur du pont sont propres à Agrionemys ( Broin 1977; Lapparent de Broin et al. 2006b, c). Toutefois chez « T. graeca terrestris Forskål, 1775 », in Gmira (1995) (dont REP 50, coll. en dépôt au MNHN.F) le bourrelet épiplastral, sans poche gulaire, s’arrête en avant de l’entoplastron; ce spécimen de Testudo est alors morphologiquement proche sur ce seul point des Chersine quaternaires et d’ Agrionemys , sa charnière hypo-xiphiplastrale étant présente pour le distinguer, entre autres différences. D’autres caractères diagnostiquent T. kenitrensis et T. oughlamensis n. sp. mais comme leur présence se manifeste éventuellement chez d’autres spécimens pliocènes de Testudo des environs d’Ahl Al Oughlam dépourvus de l’apomorphie du bourrelet concave, ils peuvent caractériser un groupe kenitrensis plus large que le clade:
1) L’encoche nucale trapézoïdale profonde du bord antérieur de la dossière, visible au moins dorsalement, avec nucale en retrait par rapport aux premières périphériques. Si les bords médiaux des périphériques 1 sont encochés, concaves, sur les paratypes de T. oughlamensis n. sp., ils apparaissent plus rectilignes chez l’holotype comme chez T. kenitrensis ; mais comme ils sont un peu érodés au bord, on ne saurait affirmer qu’ils n’aient pas été également un peu encochés. Une encoche trapézoïdale semblable à celle du clade kenitrensis , forte et visible dorsalement, existe chez le spécimen de Testudo sp. B d’Ar Rehamna décrit ci-dessous et permet de l’associer dans le groupe kenitrensis . Cette encoche nucale trapézoïdale du bord antérieur de la dossière osseuse dans le clade kenitrensis est l’accentuation profonde de la légère concavité du bord antérieur de la carapace dorsale, souvent présente chez les autres espèces de Testudo y compris T. graeca (spécimens MNHN.F en provenance d’Oran [ Algérie] REP 30, ou de Rabat [ Maroc] REP 20, par exemple); elle est visible chez les spécimens actuels quand les écailles sont enlevées pour montrer le bord osseux, lequel ne correspond pas toujours avec le bord écailleux (croissance différentielle de l’os dermique et de l’écaille sur les pourtours). Cette concavité très modérée du bord antérieur est d’ailleurs présente chez certains spécimens de Testudo graeca du Pléistocène supérieur du Maroc: Tensiftien de la petite dune de Rabat à Temara ( Gmira 1995: pl. VIB) par exemple. Une faible encoche nucale trapézoïdale très large, non visible dorsalement, existe chez les actuels comme le spécimen de T. terrestris cité ci-dessus (MNHN.F, REP 50) et chez un spécimen de T. ibera Pallas, 1814 de Turquie (REP 73) notamment. Une forte encoche nucale visible dorsalement existe, mais en V profond dont les branches partent rectilignes de chaque côté de la cervicale, chez T. kleinmanni et chez Testudo sp. du Pliocène de Çalta ( Turquie) ( Rage & Sen 1976) et de Grèce ( Garcia et al. 2011) avec un allongement très fort des périphériques encadrantes 1 et 2. Il y a donc une tendance à l’approfondissement de l’encoche du bord antérieur chez Testudo , faible au départ et soit en trapèze (à son maximum dans le groupe kenitrensis ), soit en V (à son maximum dans l’espèce de Çalta). L’encoche trapézoïdale du bord antérieur de la carapace du clade T. kenitrensis - T. oughlamensis n. sp. rappelle les actuels Kinixys et Homopus d’Afrique (carapace osseuse), avec la cervicale saillante ou non suivant les espèces comme ici dans le groupe marocain; elle rappelle aussi le jeune de Namibchersus namaquensis , l’encoche étant plus large antérieurement et s’amoindrissant chez l’adulte ( Lapparent de Broin 2003, 2008). Elle rappelle aussi « Ergilemys » bruneti Broin, 1977 de l’Oligocène de France, avec aussi des pointes périphériques à la jonction des marginales et une saillie médiane de la nucale, toutefois moins en retrait des périphériques 1. En tout état de cause, les autres écailles et plaques du groupe kenitrensis n’ont pas les mêmes proportions chez Kinixys , Homopus , Namibchersus et « E. » bruneti.
2) La forme de la vertébrale 1 qui peut être en forme de lyre chez plusieurs individus: deux spécimens de T. kenitrensis et l’holotype de T. oughlamensis n. sp., au moins. Tous les spécimens de T. oughlamensis n. sp. n’ont pas la vertébrale 1 en lyre comme ceux de T. kenitrensis . Comme il n’y a que deux spécimens connus de T. kenitrensis , la constance de la lyre au sein de cette espèce n’est peut-être pas absolue. Quoiqu’il en soit ce sont les seules espèces nommées présentant éventuellement une telle lyre en commun avec les autres caractères chez Testudo et, constant ou non, le caractère peut faire partie de la diagnose d’un groupe kenitrensis plus large que le clade; en effet, le spécimen de Testudo sp. B d’Ar Rehamna décrit ci-dessous a aussi une vertébrale 1 presque en lyre; d’autre part, un spécimen de Testudo sp. A , de Dehar Mouak décrit ci-dessous, du Pliocène un peu inférieur à celui de T. oughlamensis n. sp., possède également une vertébrale 1 en lyre, de sorte qu’il est rapproché du groupe T. kenitrensis par ce seul caractère. Chez les autres espèces de Testudo , la vertébrale 1 a les bords latéraux arrondis (le plus souvent chez les T. graeca du Maroc et d’Algérie comme souvent chez T. ibera ) sans le net resserrement assez long pour caractériser la forme de lyre (coll. REP, en dépôt au MNHN.F); ou bien ces espèces ont des bords rectilignes comme chez un spécimen d’Ahl al Oughlam ( Fig. 4 View FIG ) et, souvent, chez les jeunes spécimens actuels de Testudo , atteignant les bords latéraux de la nucale ou recoupant plus ou moins ces bords comme les autres spécimens d’Ahl al Oughlam ( Fig. 4 View FIG ); ou bien les bords latéraux de la vertébrale 1 dépassent largement la nucale, comme chez un spécimen au moins de T. cyrenaica Pieh & Perälä, 2002 (MNHN.F, REP 24).
3) La forme, large pour la longueur, courte et bien arrondie, de la dossière de T. kenitrensis et T. oughlamensis n. sp. (non totalement conservée à l’arrière chez l’holotype mais complétée par les paratypes). Les parties antérieure et postérieure de la dossière sont inclinées vers le bas ( Fig. 2A 4 View FIG ; Gmira 1995: pl. II) sans un long plateau horizontal entre elles comme il y en a chez les autres espèces de Testudo dont T. graeca ( Fig. 6C View FIG ; Gmira 1995: pls VIIB, IXC) et la petite espèce T. kleinmanni . Il en est de même pour le spécimen de Testudo sp. B d’Ar Rehamna et le caractère est aussi attribué de ce fait au groupe kenitrensis .
4) Les vertébrales sont moins larges que chez T. graeca dans les deux espèces, d’après le rapport largeur sur longueur de la vertébrale 3, de 142 % chez T. oughlamensis n. sp. et de 138 % chez T. kenitrensis pour c. 152 à 155 % chez T. graeca , mâles et femelles; la série vertébrale est encore plus étroite dans l’espèce Testudo sp. B d’Ar Rehamna (rapport de 114 % pour la vertébrale 3) rapportée de ce fait aussi au groupe kenitrensis .
T. kenitrensis et T. oughlamensis n. sp. diffèrent en outre de T. graeca par des anales plus longues médialement que chez T. graeca , pointant vers la suture hypo-xiphiplastrale de sorte que médialement leur longueur est nettement bien plus grande que la longueur des fémorales, comme par ailleurs chez T. kleinmanni et T. cyrenaica . Ce caractère n’étant pas connu chez l’espèce Testudo sp. B d’Ar Rehamna est susceptible de caractériser tout le groupe en conjonction avec les autres caractères.
T. oughlamensis n. sp. et T. kenitrensis diffèrent l’un de l’autre par:
1) le bord antérieur de la nucale (préservée chez cinq spécimens) avec une saillie médiale chez T. oughlamensis n. sp.: la saillie affecte toute la plaque médialement et pas seulement la partie recouverte par l’écaille cervicale ( 5 spécimens); la plaque a son bord antérieur rectiligne chez T. kenitrensis (deux spécimens);
2) le bourrelet dorsal moins long sur l’entoplastron chez T. oughlamensis n. sp. ( Figs 2; 4 View FIG ; 5) que chez T. kenitrensis ( Fig. 6B View FIG ), ce qui est vrai pour les 8 lots d’épiplastrons (les 3 paires des trois carapaces et 5 isolés) de T. oughlamensis n. sp. En revanche chez T. oughlamensis n. sp. le lobe antérieur est relativement plus long: les épiplastrons qui forment le bourrelet sont un peu allongés vers l’avant médialement, avec des gulaires relativement un peu plus étroites pour leur longueur;
3) l’insertion du contrefort hypoplastral, mais c’est un caractère potentiellement variable intraspécifiquement; elle se fait principalement sous la périphérique 7 avec un petit débordement ventral sous la périphérique 8 chez T. oughlamensis n. sp. (comme chez un T. ibera de Turquie et chez certains T. marginata de Grèce); alors qu’elle se fait complètement entre les deux plaques chez T. kenitrensis (visible sur l’holotype seul) comme généralement chez Testudo , où l’insertion peut aussi se faire: sous la 7 e seule (chez certains T. graeca d’Afrique du Nord ); sous la périphérique 7 avec un petit débordement dorsal sous la périphérique 8 (chez quelques T. graeca du Maroc et T. terrestris ); et sous la 8 e principalement (chez un T. graeca d’Afrique du Nord );
4) les écailles vertébrales 2, 3 et 4, relativement plus étroites (largeur par rapport à la longueur de chaque écaille et particulièrement la 3e) distinguent l’holotype de T. kenitrensis de celui de T. oughlamensis n. sp. à vertébrales un peu plus larges; l’amplitude de la variabilité (compte tenu du sexe notamment) ne peut être établie mais ce qui reste des spécimens AaO-948 et AaO-949 ( Fig. 4 View FIG ) confirme l’holotype ( Fig. 3 View FIG ), tous trois devant être des femelles comme T. kenitrensis d’après le plastron plat;
5) il n’y a qu’une grande écaille inguinale chez T. kenitrensis au lieu de deux petites chez T.oughlamensis n. sp. (et chez Testudo graeca il y en a deux dont l’externe plus grande).


| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Testudo oughlamensis
| Gmira, Saloua, Broin, France De Lapparent De, Geraads, Denis, Lefèvre, David, Mohib, Abderrahim & Raynal, Jean-Paul 2013 |
Testudo
| BAILON S. 2000: 542 |
| LAPPARENT DE BROIN F. DE 2000: 62 |