
Mycale (Kerasemna) humilis ( Thiele, 1903 ), 2021
|
publication ID |
https://doi.org/10.11646/zootaxa.4912.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9536C1CF-4AEF-47F8-959B-48CD7A5392D8 |
|
DOI |
https://doi.org/10.5281/zenodo.4473234 |
|
persistent identifier |
https://treatment.plazi.org/id/361087A7-FFA4-FFC6-55AB-FCE753CDCA14 |
|
treatment provided by |
Plazi |
|
scientific name |
Mycale (Kerasemna) humilis ( Thiele, 1903 ) |
| status |
comb. nov. |
Mycale (Kerasemna) humilis ( Thiele, 1903) View in CoL comb.nov.
Figs 61 View FIGURE 61 a–d, 62a–f, 63a–g, 64
Biemna humilis Thiele, 1903: 944 , fig. 10.
? Desmacella tubulata sensu Dendy 1916: 116 (not: Dendy 1905).
Sigmatoxella humilis View in CoL ; Pulitzer-Finali 1982a: 107.
Mycale aff. c ockburniana ; Van Soest 1990: 305; Erhardt & Baensch 1998: 54 (not: Hentschel 1911)
? Desmacella humilis ; Pulitzer-Finali 1993: 293.
? Kerasemna humilis View in CoL ; Richmond et al. 2011: 122, fig. p. 123.
Mycale (Mycale) vansoesti Calcinai et al., 2006: 197 View in CoL , figs 3A–C, 4A–F, 5A–J, 6A–F (not: Hajdu 1995).
Mycale (Mycale) corallina Calcinai et al. 2017 View in CoL : Supplementary file 2.
Mycale calcinaiae Van Soest & Hooper, 2020: 63 View in CoL .
Mycale humilis View in CoL ; Van Soest & Hooper, 2020: 63.
Material examined. ZMB 3203, two slides of holotype of Biemna humilis (slides labeled as Desmacella humilis ), Indonesia, Ternate, coll. W. K̹kenthal, 1893–1894; MSNG 52803, paratype of Mycale (Mycale) vansoesti Calcinai et al., 2006 , Indonesia, N Sulawesi, Siladen Station, depth 17 m, 28 May 2002.
ZMA Por. 02905, Indonesia, Lesser Sunda Islands ( Nusa Tenggara), Postillion Islands, anchorage off Pulau Sarassa , 7.1833°S 118.2°E, coral bottom, depth 36 m GoogleMaps , dredge, coll. Siboga Expedition stat. 043, field nr. SE829I, 4 April 1899; ZMA Por. 08028, Indonesia, Nusa Tenggara East , Komodo, NE cape, 8.4833°S 119.5683°E, depth 1–4 m GoogleMaps , snorkeling, coll. R. W.M. van Soest, Indonesian-Dutch Snellius II Expedition stat. 096, field nr. 096 / II/02 , 19 September 1984 (live color yellow) ; ZMA Por. 08564, Indonesia, Sulawesi, SE Sulawesi, Taka Karlarang , reef, depth 6–8 m , SCUBA, coll. R. W.M. van Soest, Indonesian-Dutch Snellius II Expedition stat. 147, field nr. 147 / III/11 , 27 September 1984 (yellow) ; ZMA Por. 08953, Sulawesi, SE Sulawesi, SW Salayar, reef N of Pulau Bahuluang, 6.45°S 120.43°E, depth 10–15 m GoogleMaps , SCUBA, coll. R. W.M. van Soest, Indonesian-Dutch Snellius II Expedition stat. 169, field nr. 169 / IV/24 , 30 September 1984 (light yellow) ; ZMA Por. 09753, Papua New Guinea, no further data, dried, coll. M.C. Díaz, field nr. 90152 (orange-red) ; ZMA Por. 12970, Indonesia, N Sulawesi, Bunaken Island , 1.6132°N 124.7797°E, reef drop-off, dried, coll. H. Erhardt, 22 September 1996 (white-purple, photo in Erhardt & Baensch 1998) GoogleMaps ; ZMA Por. 13327, Indonesia, Sulawesi, SW Sulawesi, Spermonde Archipelago, Samalona , reef, depth 6 m , SCUBA, coll. N.J. de Voogd, field nr. SA/NV/060597/03, 6 May 1997 (white-purple) ; ZMA Por. 14523, Indonesia, N Sulawesi, Bunaken, ESE Siladen Island , 1.6317°S 124.8053°E, depth 18 m GoogleMaps , coll. B.W. Hoeksema, SYMBIOSPONGE Expedition, field nr. 98/NS/MAY07/BH/077, 7 May 1998 (white); ZMA Por. 15829, Madagascar, Nosy B, Crater Bay , depth 4–10 m , muddy reef bottom, SCUBA, coll. J.H. Stock, 21 December 1963 (orange) ; ZMA Por. 16832, Indonesia, Maluku, Ambon, Tanjung Setan , depth 20 m , SCUBA, coll. S. Weinberg, 20 August 1995 (orange) ; RMNH Por. 1608, Palau, Koror, Ngerikuul Pass, E of Ngetekiou Island, pass between Ngeteklou and Ngermeuangel Island , 7.3167°N 134.5167°E, depth 8 m GoogleMaps , SCUBA, coll. N.J, de Voogd, field nr . KOR02 About KOR /120505/003, 12 May 2005 (green) ; RMNH Por. 1609, Palau, Koror, Ngerikuul Pass, E of Ngetekiou Island, pass between Ngeteklou and Ngermeuangel Island , 7.3167°N 134.5167°E GoogleMaps , SCUBA, coll. N.J, de Voogd, field nr . KOR02 About KOR /210505/095, 21 May 2005 ; RMNH Por. 1610, Palau, Koror, Ngerikuul Pass, E of Ngetekiou Island, pass between Ngeteklou and Ngermeuangel Island , 7.3167°N 134.5167°E GoogleMaps , SCUBA, coll. N.J, de Voogd, field nr . KOR02 About KOR /210505/096, 21 May 2005 ; RMNH Por. 2150, Bali, N side of Nusa Penida, off Desa Buyuk , 8.6736°S 115.5436°E, depth 0–30 m GoogleMaps , SCUBA, coll. N.J. de Voogd, Bali-Lombok Strait Expedition 2001, field nr. BAL.34/210401/259, 21 April 2001 (white) ; RMNH Por. 2190, Bali, Padang Bai, E side Tanjung Sari , 8.5294°S 115.5131°E, shallow slope, depth 0–30 m GoogleMaps , SCUBA, coll. N.J. de Voogd, Bali-Lombok Strait Expedition 2001, field nr. BAL.19/080401/097, 8 April 2001 (white) ; RMNH Por. 2608, Singapore, Pulau Tekukor ( Monkey Island ), NW side, 1.2308°N 103.8365°E, depth 12 m GoogleMaps , SCUBA, coll. N.J. de Voogd, field nr. SIN.23/040406/170, 4 April 2006 (yellow) ; RMNH Por. 2617, Indonesia, Sulawesi, SW Sulawesi, Tanjung Bira, Pulau Liukan , 5.65°S 120.43°E, depth 25 m GoogleMaps , SCUBA, coll. N.J. de Voogd, field nr. BIR05/230501/242, 23 May 2001 ; RMNH Por. 2618, Indonesia, North Sulawesi, SW Siladen Island, 1.6167°N 124.7833°E, depth 15 m GoogleMaps , SCUBA. coll. N.J. de Voogd, field nr. MD11 /170502/079, 17 May 2002 ; RMNH Por. 3054, Indonesia, Sulawesi, SW Sulawesi, Tanjung Bira, Pulau Liukan , 5.65°S 120.43°E, depth 28 m GoogleMaps , SCUBA, coll. N.J. de Voogd, field nr. BIR05/230501/321, 23 May 2001 ; RMNH Por. 5329, Indonesia, Halmahera , Maitara Maitara W, 0.7299°N 127.3624°E, depth 12 m GoogleMaps , SCUBA, coll. N.J. de Voogd, Ternate-Halmahera Expedition 2009, field nr. TER.09/291009/080, 29 October 2009 ; RMNH 8867 About RMNH , Taiwan, Dong YiJu , west side, outside fishing harbor, 23.2552°N 119.6671°E GoogleMaps , SCUBA, coll. N.J. de Voogd, field nr. PES135, 28 July 2014 (orange) .
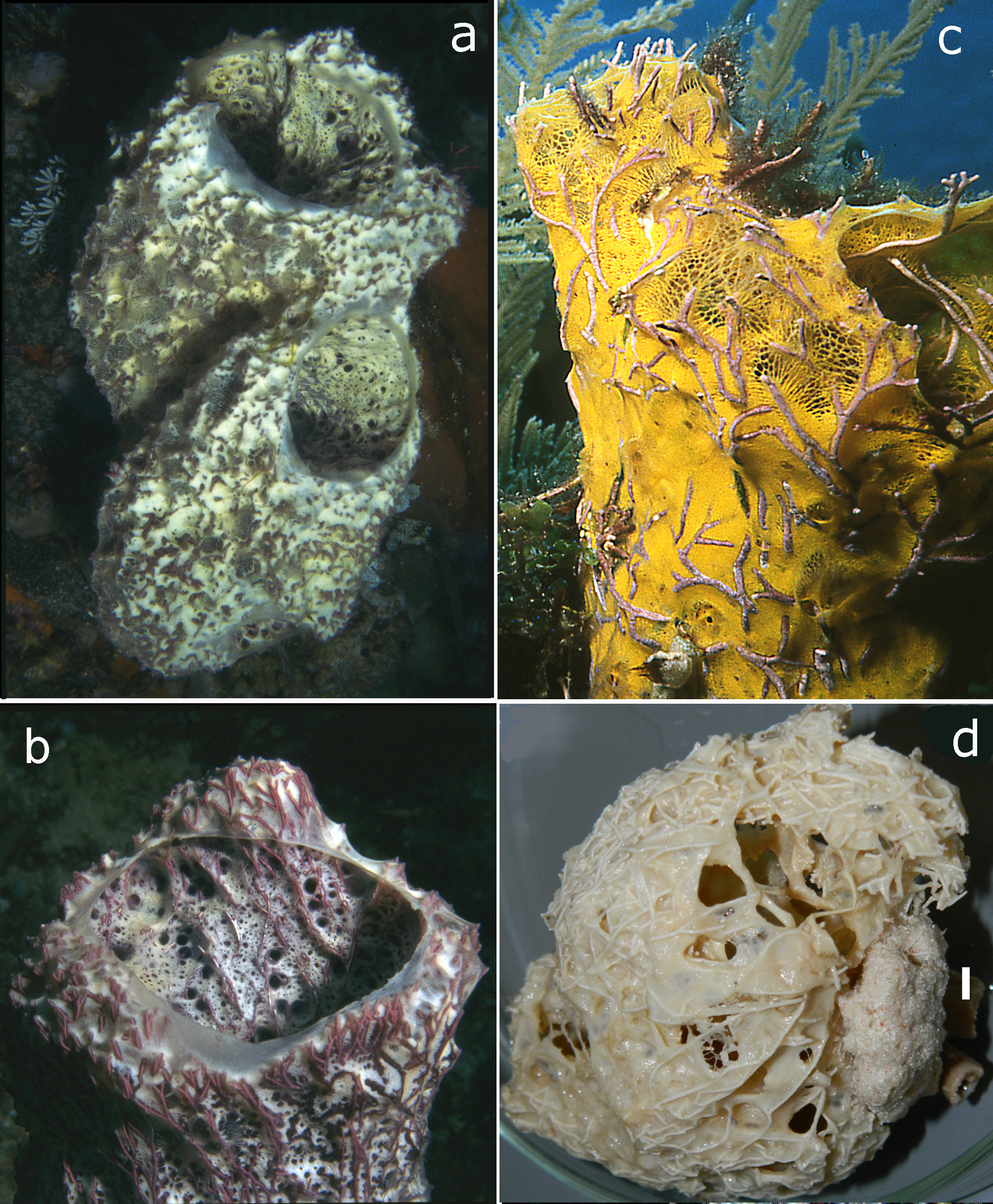
Description ( Figs 61 View FIGURE 61 a–d, 63a). The species was recently described extensively by Calcinai et al. (2006) as Mycale (Mycale) vansoesti . We can confirm their excellent account of the species, also based on slides made from a fragment of the paratype sent to us by Dr. Barbara Calcinai. The specimens in their full-grown habitus exhibit wide-mouthed tubes, often two such tubes are found merged to a larger mass with two openings. Size may be considerable: Calcinai et al. (2006) report specimens of 40 cm high and 20 cm wide, with terminal vent up to 15 or 20 cm. The walls of the tube are thin (often less than 1 cm in thickness), web-like, supported by purple- or red-colored algal strands (genus Amphiroa ). Juvenile or incipient growth forms may be much smaller and basically are encrusting. These may lack an upper large opening, in stead having lateral excurrent openings giving access to the hollow interior. The surface is irregularly conulose, caused by Amphiroa strands lifting up the surface tent-like. The skin between the strands, both on the outside and the inside of the tubes is punctured by many small incurrent and excurrent openings, of about 1 mm or less in diameter. The inner surface of the vent has also irregularly distributed larger openings, presumably oscules. Colors of larger specimens sometimes give an overall pinkish impression ( Figs 61 View FIGURE 61 a–b), due mostly to the algal strands, but the tissue inbetween is pale yellow, and yellow-orange colors ( Fig. 61c View FIGURE 61 ) are also reported frequently, especially in smaller encrusting specimens, occasionally white, rarely greenish. So far the colors are not clearly correlated to other features of the species, so we assume them to be variable. Preserved, colors are off-white or light beige ( Figs 61d View FIGURE 61 , 63a View FIGURE 63 ). Consistency is fragile, easily damaged.
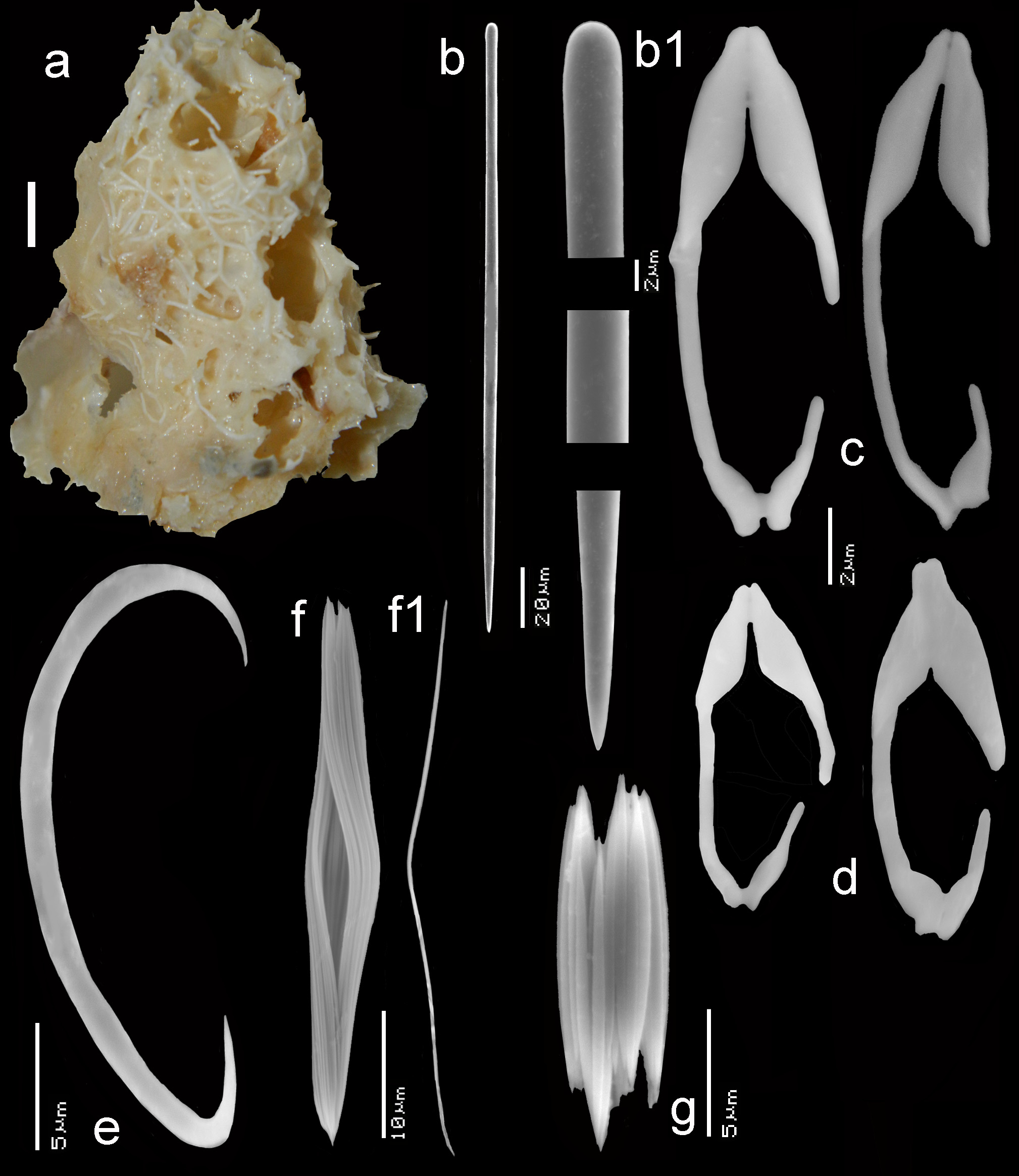
Skeleton ( Figs 62 View FIGURE 62 a–f). The algal strands (thickness 250–400 µm) form the framework of the skeleton, but spongin-encased spicule tracts of variable thickness (commonly 30–70 µm in diameter, occasionally as thick as 200 µm) and length (may be as long as 1 mm or more) connect the algal strands to form a secondary supporting structure. Between the algal strands and the spicule tracts there is an unorganized mass of toxodragmas, anisochelae, sigmas, rare trichodragmas and single tangentially arranged megascleres. No clear ectosomal skeleton on both sides of the tube walls. The toxodragmas may form vague clusters, but sigmas and anisochelae are scattered singly in the tissue, no rosettes.
Spicules ( Figs 63 View FIGURE 63 b–g). Mycalostyles, two categories of anisochelae, sigmas, toxas (mostly in toxodragmas), trichodragmas.
Mycalostyles ( Fig. 63b,b View FIGURE 63 1 View FIGURE 1 ), comparatively short and robust, with barely developed head, slightly fusiform, 207– 242.7 –297 x 4– 6.7 – 11 µm; type specimen: 231– 267.3 –297 x 4.5– 7.4 – 11 µm.
Anisochelae I ( Figs 63c View FIGURE 63 ), usually less common, narrow-shaped but otherwise normal, with both upper and lower alae developed, free part of the shaft about 40–50 %, size 17– 22.9 – 30 µm; type specimen: 17– 19.4 – 22 µm.
Anisochelae II ( Figs 63d View FIGURE 63 ), usually more common, comparatively slightly more reduced with upper and lower alae pointed towards each other, 10– 13.4 – 19 µm; type specimen: 10– 13.3 – 16 µm.
Sigmas ( Fig. 63e View FIGURE 63 ), usually very common, thin, asymmetrically curved, the endings look roughened under SEM, but no definite spines were observed, 16– 24.9 – 30 µm; type specimen: 19– 24.8 – 30 µm.
Toxodragmas ( Figs 63f,f View FIGURE 63 1 View FIGURE 1 ), extremely common, consisting of an intertwined double-pack of individual thin raphide-like toxas (less than 1 µm in thickness), leaving a characteristic eye-like central opening, 33– 42.5 –51 x 4– 6.6 – 10 µm; type specimen: 34– 43.9 –51 x 4– 7.4 – 10 µm.
Trichodragmas ( Fig. 63g View FIGURE 63 ), not common, consisting of a small package of straight raphides, 7– 11.8 –17 x 3– 6.6 – 10 µm; type specimen: 9– 11.9 –15 x 3– 6.4 – 9 µm.
Distribution and ecology ( Fig. 64 View FIGURE 64 ). Indonesia, Singapore, Papua New Guinea, Palau, Taiwan, Madagascar, Kenya,?Western India, on reefs down to 36 m. More ecological information is provided in Calcinai et al. (2006).
Remarks. We re-examined original slides of Thiele’s Biemna humilis and found abundant anisochelae I and II, apparently overlooked by Thiele. In most aspects, Biemna humilis conforms with what is so far known as Mycale aff. cockburniana sensu Van Soest 1990 and Mycale (Mycale) vansoesti Calcinai et al., 2006 . Thiele’s material was a blade-like encrustation, not an elaborate wide-tubed specimen, but in spiculation and presence of Amphiroa strands there is complete conformity. Calcinai et al. ’s name is a primary junior homonym of Mycale (Grapelia) vansoesti Hajdu, 1995 , which has priority. Calcinai et al. 2017 proposed Mycale (Mycale) corallina to replace M. (M.) vansoesti Calcinai et al. , because junior primary homonyms are permanently invalid (ICZN Art. 57.2). Van Soest & Hooper (2020: 63) were unaware of this replacement name (which was proposed in a Supplementary file, and at the time was not picked up by the World Porifera Database) and proposed Mycale calcinaiae as a replacement name. Both names are junior synonyms of Mycale (Kerasemna) humilis ( Thiele, 1903) comb.nov.
Calcinai et al.’s description does not entirely conform to our above description as it omitted reporting the small trichodragmas, which were found in all our specimens, including Calcinai’s paratype. Because they are quite small and not at all very common, we assume that they were overlooked by Calcinai et al.
Although Pulitzer-Finali (1993) reported the present species from Kenya as Desmacella humilis , he pointed out in the discussion that it shared the toxodragmas with his Australian Kerasemna tenuityla . Subsequently, Kelly in Richmond et al. 2013, used the combination Kerasemna humilis for our species. We confirm here the presence of the species in the Western Indian Ocean by reporting a specimen from Madagascar. The spicule shape and sizes (mycalostyles 244–249 x 6–7.5 µm, anisochelae I 22–25 µm, anisochelae II 17–19 µm, sigmas 21–29 µm, toxodragmas 45–50 µm, and trichodragmas 12–17 µm) conform to those of the numerous specimens we report from the Western Pacific. Apparently orange colored specimens are more common there, as both our specimen, Pulitzer-Finali’s specimens and Richmond’s specimen are reported to have that color. Still, orange and yellow specimens were also reported from Indonesia (see Fig. 61c View FIGURE 61 ), Singapore, Taiwan and Papua New Guinea. The coralline algae found associated with the species recognized so far appear to be mostly Amphiroa spec., at least in the pink, purple, or pale yellow-white forms. Possibly, the association is thus obligatory, but this may be only for these forms. Yellow-orange specimens may have a different algal associate (cf. Calcinai et al. 2006). Pulitzer-Finali’s (1993) Kenya and Zanzibar records mention the presence of toxodragmas and sigmas, but do not report anisochelae. It is possible that the specimens do not contain anisochelae and belong to M. (K.) tenuiyla rather than to M. (K.) humilis .
Dendy’s (1916) report of a deviating Desmacella tubulata appears on paper to be a likely record of the present species from Western India, as Dendy mentions ‘toxa arranged in a sheaf or toxodragma’. This will need to be reexamined for certainty.
Mycale cockburniana Hentschel, 1911 and its variety albanensis Hentschel, 1911 , generally shares the spicule complement with the present species: mycalostyles 192–256 x 3–4 µm, two size categories of anisochelae 21–27 µm and 13–16 µm, thin sigmas 22–25 µm, and trichodragmas 25–30 µm. The habitus is encrusting and contains ingrown ‘Pflanzentheile’, adding to the similarity. The main difference is the absence of the peculiar toxodragmas. The skeleton is described as conforming to subgenus Carmia (cf. above).
Kerasemna tenuityla Pulitzer-Finali, 1982 (see below) is a close relative, differing mostly in the absence of anisochelae. We maintain this as a separate species for the time being.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Mycale |
Mycale (Kerasemna) humilis ( Thiele, 1903 )
| Van, Rob W. M., Aryasari, Ratih & De, Nicole J. 2021 |
Kerasemna humilis
| Richmond, M. D. 2011: 122 |
Mycale (Mycale) vansoesti
| Calcinai, B. & Cerrano, C. & Totti, C. & Romagnoli, T. & Bavestrello, G. 2006: 197 |
Desmacella humilis
| Pulitzer-Finali, G. 1993: 293 |
Mycale aff. c ockburniana
| Erhardt, H. & Baensch, H. A. 1998: 54 |
| Van Soest, R. W. M. 1990: 305 |
Sigmatoxella humilis
| Pulitzer-Finali, G. 1982: 107 |
Desmacella tubulata sensu Dendy 1916: 116
| Dendy, A. 1916: 116 |
Biemna humilis
| Thiele, J. 1903: 944 |