
Thelastoma vanwaerebekei, Orsini & Cuellar & Rondan Dueñas & Gardenal & Doucet & Lax, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4375.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:0CDB3909-22C2-4096-B351-8587082A70B1 |
|
DOI |
https://doi.org/10.5281/zenodo.5967971 |
|
persistent identifier |
https://treatment.plazi.org/id/2D0887C7-FF96-271D-3B8F-81FBFD29FA46 |
|
treatment provided by |
Plazi |
|
scientific name |
Thelastoma vanwaerebekei |
| status |
sp. nov. |
Thelastoma vanwaerebekei View in CoL n. sp.
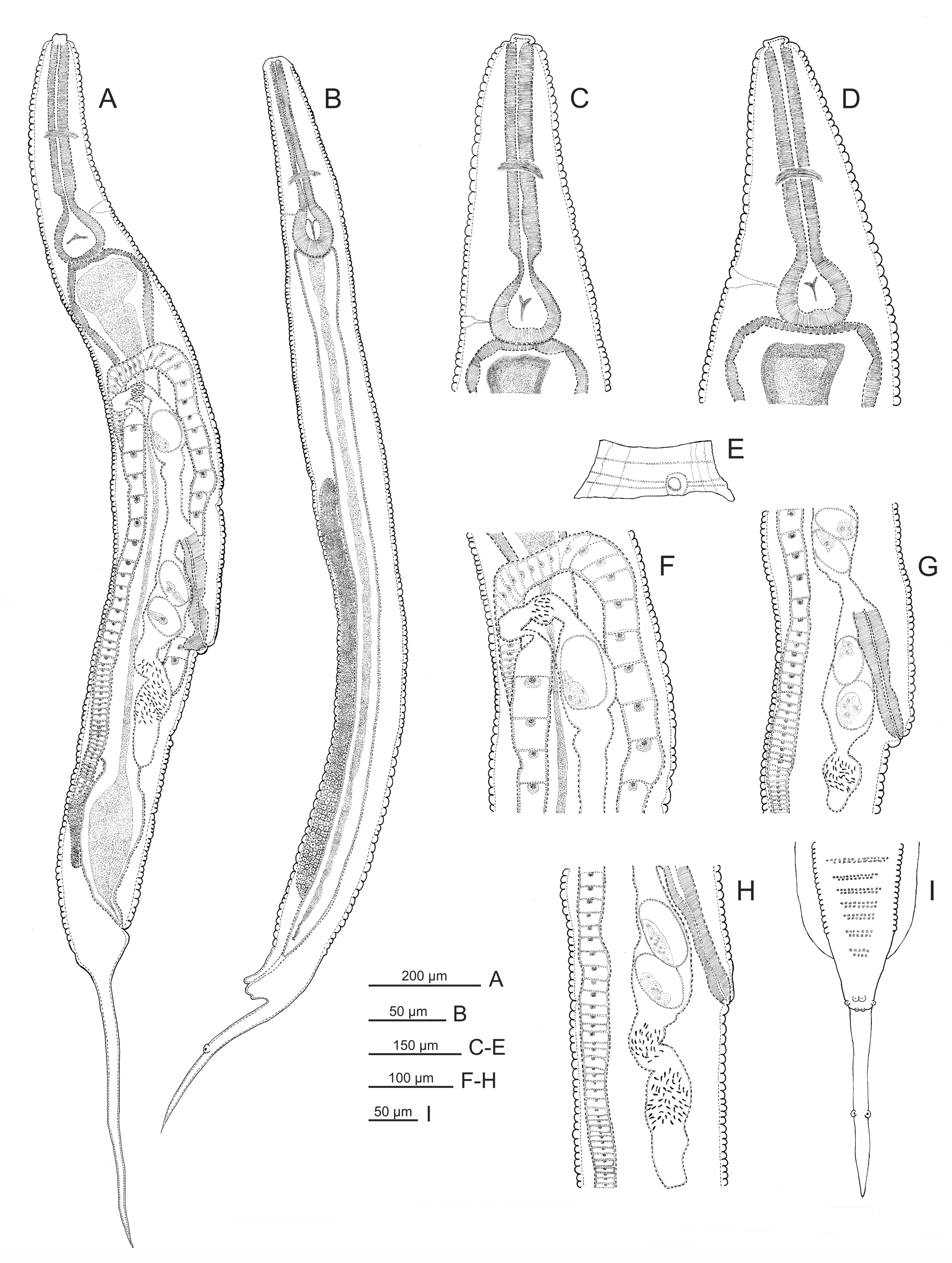
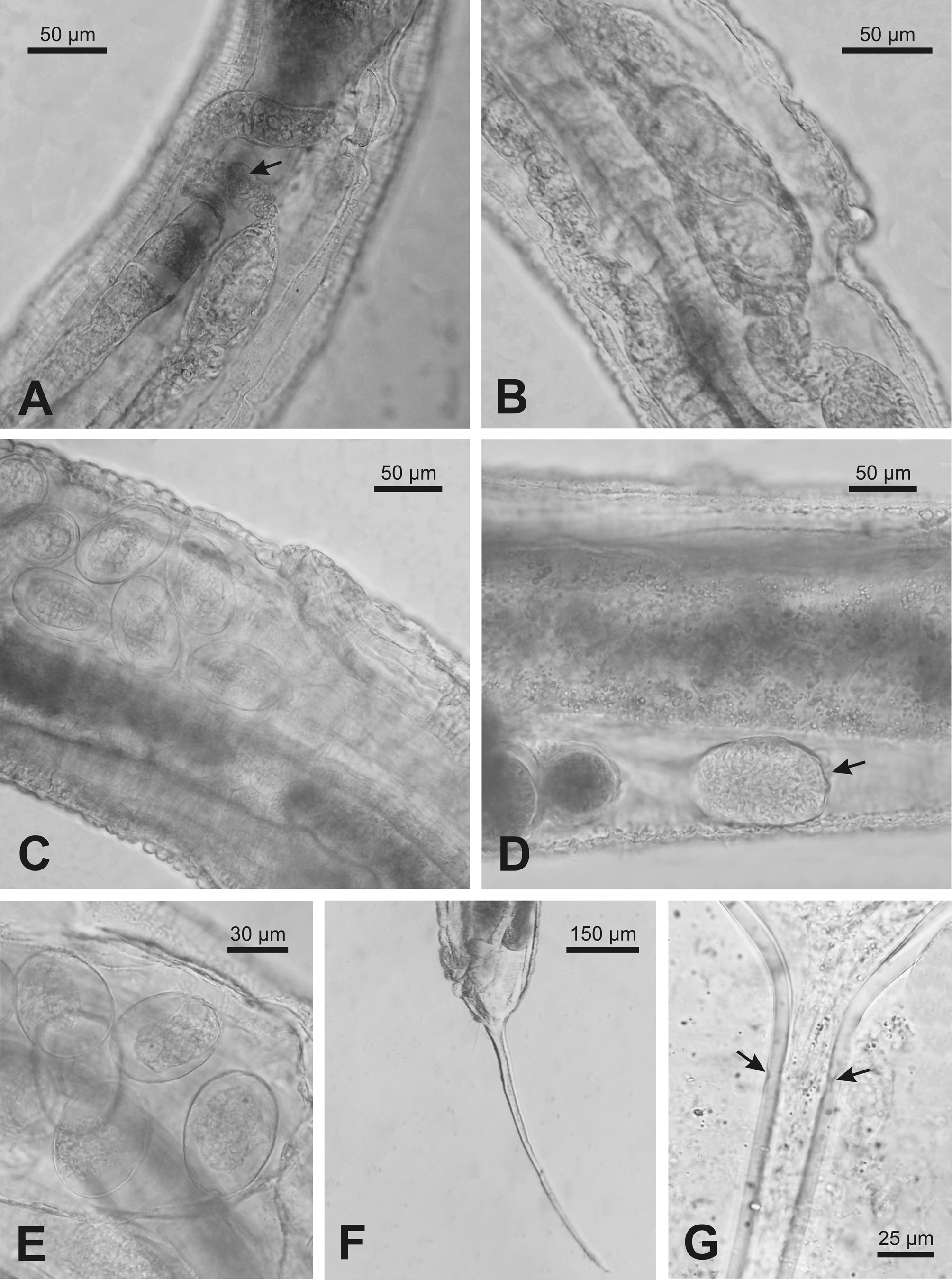
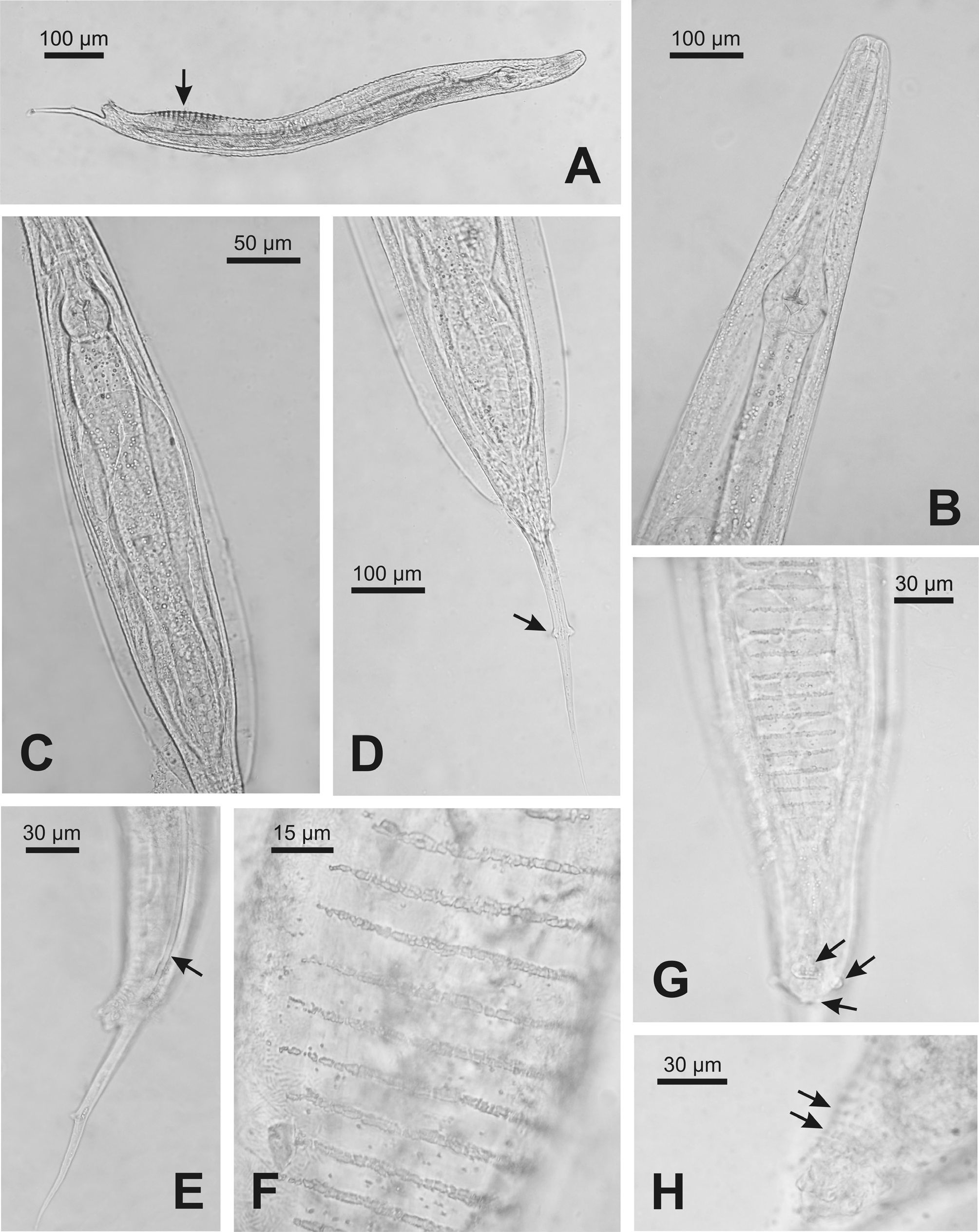
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE3 View FIGURE 4 )
Measurements. See Table 1.
Description (based on 30 females and males, including the holotype and allotype).
Female. Body spindle-shaped, reaching maximum width at the vulval region. Cuticle transversally striated. Lack of lateral alae. Labial annule clearly set off from the rest of the body; first body annule large, extending to the beginning of the pharynx corpus. Posteriorly, annules more reduced in size and become almost undetectable behind the anus. Buccal cavity rectangular, with sclerotized walls. Pharynx composed of three parts: corpus, isthmus and basal bulb. Nerve ring located approximately at the middle region of pharynx. Corpus cylindrical, set off from isthmus by a more or less marked constriction. Basal bulb ovoid, with refringent valvular apparatus. Excretory pore located at 15.2% (10.3–19.2%) of total body length and situated at level of mid-basal bulb in most of the specimens; it was observed near the beginning of the basal bulb only in 3 individuals. Cardia present. Intestine dilated at anterior end, followed by a gradual narrowing toward posterior end. Reproductive system didelphicamphidelphic. Vulval slit located approximately at midbody; vulval lips protuberant in 47% of the analyzed specimens, being slightly protruding in the remaining (53%) individuals. Vagina anteriorly directed; each one of the uteri is connected with an ovary. One ovary extends posteriorly, flexing between the vulva and the anus and then directing anteriorad again; it curves near the anterior portion of intestine, ending at level of vulva. The other ovary extends towards the anterior region, flexing near the beginning of the intestine, and ending near the anus. A seminal receptacle with sperm is observed in each of the branches of the reproductive system, between the uterus and the ovary. In the branch directing posteriorly, the seminal receptacle is easily visible and is situated ventrally between the vulva and the anus; in the anteriorly extending branch, the seminal receptacle is located approximately at level of the flexing, being difficult to distinguish in gravid females. Uterus containing numerous oval eggs; egg surface is smooth in vivo and irregular in fixed specimens. Posteriorly to the nematode body, the intestine becomes tubular and in the posterior portion it connects with the rectum, which ends in the anus. Tail filiform, representing 28.2% (21.7–33.3%) of total body length. Phasmids are located laterally at the beginning of caudal filament.
Male. Smaller than females. Body thin, truncate at level of anus, ending in a filiform caudal appendage. Cuticle with regular transverse striations with annules increasing gradually and becoming more pronounced in the mid region of body, where it reaches its maximum width. Pair of longitudinal lateral alae; they extend approximately from the level of the nerve ring in the isthmus, gradually widening and ending anteriorly to anus. Labial ring barely differentiated from the rest of the body. Buccal cavity rectangular; pharynx composed of a cylindrical corpus that narrows slightly to connect with a short cylindrical isthmus. Basal bulb ovoid with a refringent valvular apparatus. Nerve ring located approximately at 60% of pharynx length. Excretory pore located at 12.8% (8.6–15.2%) of total body length, at level of mid-basal bulb. Intestine tubular, simple. Testis long; it extends from the cloacal aperture, folding on itself. In vivo, anteriorly to anus and in the ventral region, a series of thin transverse lines composed of small protuberances were observed in the cuticle; these lines were highly variable in length and number. Another group of similar lines was distinguished at midbody in some individuals. In addition, irregularly arranged protuberances were observed near the anus; these structures were not observed in fixed and transparent specimens. Four pairs of caudal papillae: one prominent ventral preanal pair, overlapping the anus; one adanal pair, one postanal ventral pair, and one pair, located approximately at mid caudal filament. Lack of spicule and gubernaculum. Tail long, filiform, representing 15.3% (11.2–19.6%) of total body length.
* At the vulva level (females).
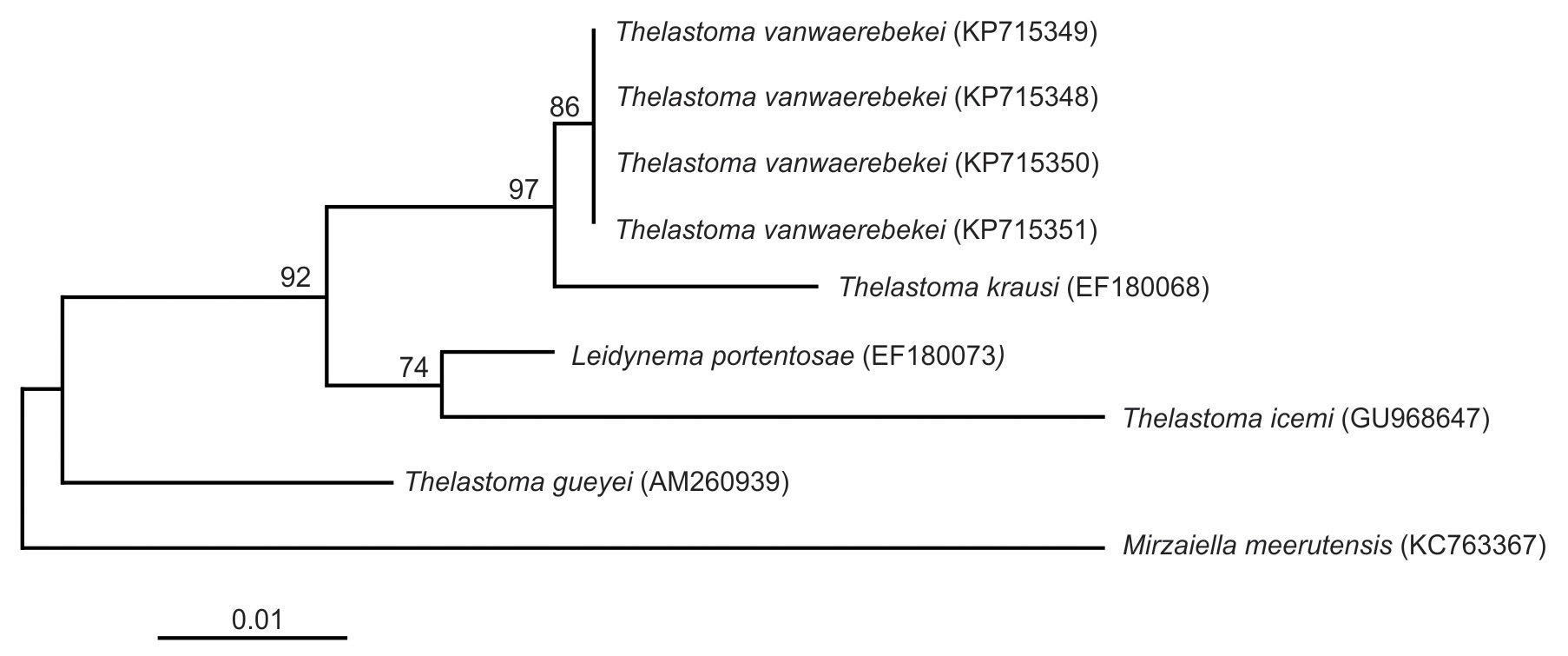
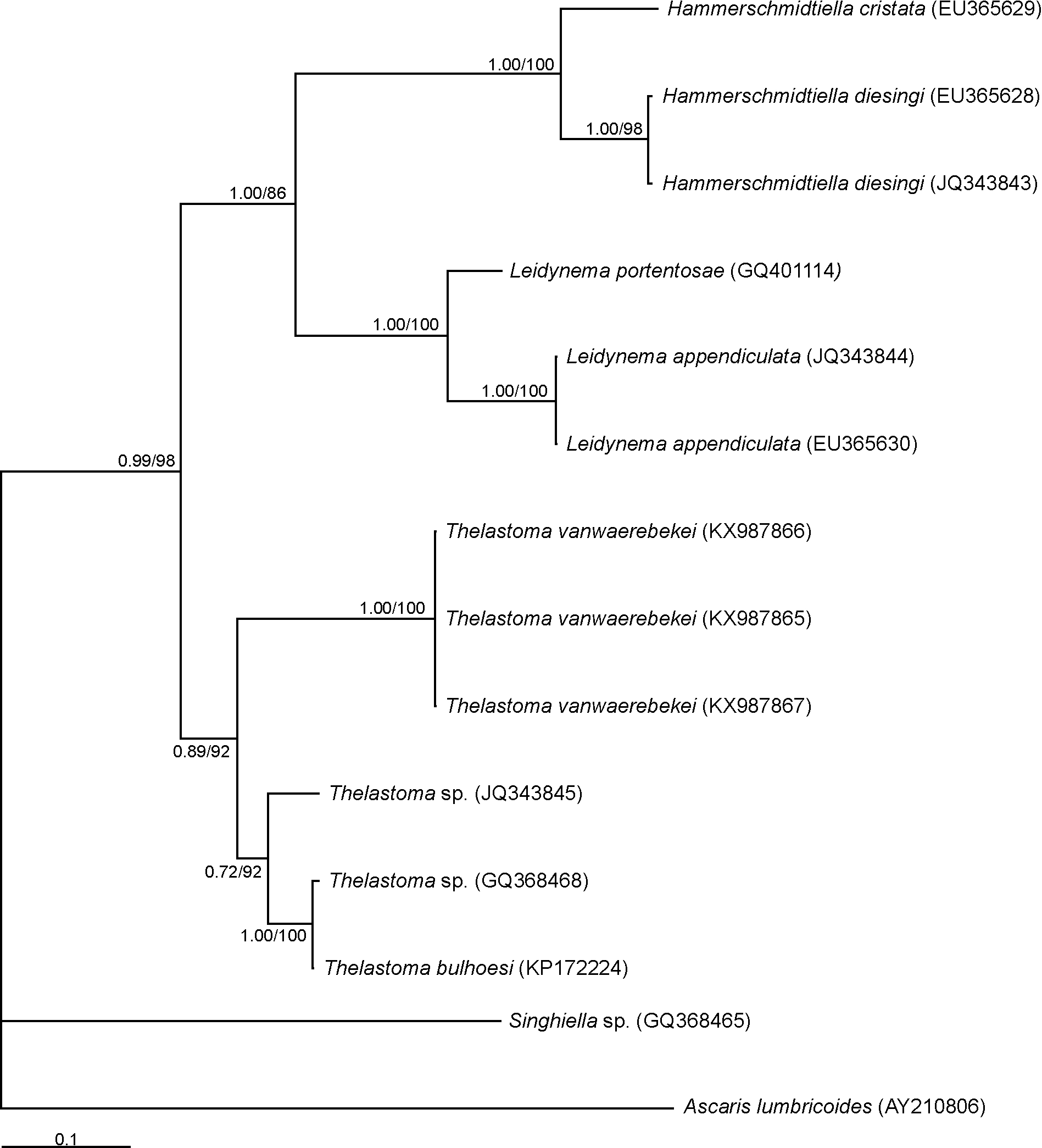
Molecular characterization and phylogenetic analysis. Thelastoma vanwaerebekei n. sp. was characterized genetically by the sequences of the 18S SSU rRNA ( KP715348 View Materials – KP715351 View Materials ) and the D2-D3 expansion segments ( KX987865 View Materials – KX987867 View Materials ). A fragment of 893 bp was sequenced for the 18S gene region in the new species; the genetic similarity percentages with the available Thelastoma sequences were 98% with T. krausi Carreno, 2007 and 95% with T. icemi and T. gueyei Koubková, Baruš, Matějusová, Hodová & Koubek, 2006 . The phylogenetic relationship of T. vanwaerebekei n. sp. with sequences of the other Thelastoma spp. inferred from 18S sequences using the ML method, is shown in Figure 5 View FIGURE 5 . The analysis showed a well-supported group (97% bootstrap) that comprised T. vanwaerebekei n. sp. and T. krausi . This group was closer to the clade (74% bootstrap) comprising T. icemi and Leidynema portentosae van Waerebeke, 1978 . Thelastoma gueyei showed a marked separation from the rest of Thelastoma spp.
For the D2-D3 regions, a fragment of 752 bp was sequenced. The genetic similarity percentages obtained were 85% with two unidentified species of Thelastoma ( JQ343845 View Materials , GQ368468 View Materials ) and T. bulhoesi ( KP172224 View Materials ). Phylogenetic tree inferred by BI showed that T. vanwaerebekei n. sp. formed a clade with the known sequences of T. bulhoesi and Thelastoma sp.. This group had a close relationship with Leidynema spp. and Hammerschmidtiella spp. ( Fig. 6 View FIGURE 6 ); the same tree topology was observed with the ML method (data not shown).
Taxonomic summary
Type host and origin: Larvae of Gymnetis litigiosa ( Coleoptera : Scarabaeidae ) from Uruguay (locality unknown).
Site of infection: hindgut.
Ecological parameters: Prevalence: 74%; mean intensity: 11.9 nematodes per coleopteran infected larva (n=23). Not found in adults of the insect (n=20).
Type material: Holotype female (CZA TVWF 01); allotype male (CZA TVWM 01); paratype 29 females (CZA TVWF 02–30), and paratype 29 males (CZA TVWM 02–30) are deposited in the Laboratorio de Nematología [Instituto de Diversidad y Ecología Animal (CONICET-UNC) and Centro de Zoología Aplicada, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. Córdoba, Argentina].
Voucher DNA sequences: 18S SSU rRNA gene, partial sequence, GenBank accession numbers KP715348 View Materials – KP715351 View Materials . The 28S LSU rRNA gene, partial sequence, GenBank accession numbers KX987865 View Materials – KX987867 View Materials .
Etymology: This species is named in honor of the Belgian parasitologist Daniel van Waerebeke who was a specialist of nematodes of arthropods. He described several species of the Thelastomatidae family.
Diagnosis and relationships. The vulva of females of Thelastoma vanwaerebekei n. sp. is located approximately at midbody, with a seminal receptacle on each reproductive branch. The excretory pore is located at the level of the mid-basal bulb or slightly near the isthmus. The tail is filiform, representing 28% of body length. Males have lateral alae extending approximately from the beginning of the pharynx basal bulb, gradually widening and ending anteriorly to the anus. Presence of 4 pairs of caudal papillae: one prominent ventral preanal pair, one adanal pair, one postanal ventral pair and a pair located approximately at mid caudal filament. Spicule and gubernaculum absent.
On the basis of the ratio of female tail length to the total body length (in %), T. vanwaerebekei n. sp. belongs to group “C” of Thelastoma species defined by Baruš & Koubková (2002). This group comprises species with a medium to long, filiform or subfiliform tail, representing 12–30% of the total body length. It is the largest group since it comprises 30 species and two populations identified as Thelastoma sp.; it includes species with unknown males and species with males with/without spicules. Thelastoma vanwaerebekei n. sp. is within the group of species with males lacking spicules and females having their excretory pore at the same level as the bulb (or slightly anteriorly). Thelastoma vanwaerebekei n. sp. differs from T. raoi ( Rao & Rao, 1966) Adamson & van Waerebeke, 1992 in the number of caudal papillae (4 vs. 3 pairs). Females of T. vanwaerebekei n. sp. differ from those of T. madecassum van Waerebeke, 1969 and Thelastoma sp. B ( Dollfus, 1964) in tail shape (filiform vs. conical), and from T. depressum ( Hammerschmidt, 1838) Leidy, 1853 in pharynx length (approximately 17% of body length vs. one-fourth to one-third of the body length). Thelastoma vanwaerebekei n. sp. differs from T. blabericola Leibersperger, 1960 in tail length of males (139.2–232.1 µm vs. 290–370 µm). Thelastoma sp. ( Spiridonov & Ivanova 1998) and T. basiri Farooqui, 1970 have a hood-like cephalic expansion, which is also the first body annule; this characteristic is not present in T. vanwaerebekei n. sp. Thelastoma imphalensis Chinglenkhomba, Gambhir, Tarnita, Gyaneswori & Indrani, 2007 is characterized by having a first annule well developed and distinctly set off from the rest of the body annules defining a cephalic expansion. Thelastoma dessetae Adamson, 1985 has the body cuticle anterior to the first annulation, forming a truncate cone which extends well posterior to beginning of the pharynx. None of those characteristics mentioned for the cephalic region of T. imphalensis and T. dessetae were observed in T. vanwaerebekei n. sp.
Discussion
Thelastoma vanwaerebekei n. sp. is the first species of the genus found in larvae of G. litigiosa , thus increasing the number of species with a coleopteran host: T. depressum , T. figuli van Waerebeke, 1970 , T. gallicum Théodoridès, 1955 , T. macramphidium Christie, 1931 , T. modestus , T. patellae van Waerebeke, 1970 , T. pterygoton Poinar, 1973 , T. pyrrhus van Waerebeke 1973 , T. raoi , T. rara , T. ritteri van Waerebeke, 1973 , T. robustum Leidy, 1850 , T. tetradymos , T. toxi van Waerebeke, 1970 and T. unicoloris van Waerebeke, 1970 .
Females of T. vanwaerebekei n. sp. exhibited variability in some of the evaluated characters, such as: pharynx (more or less marked corpus-isthmus junction), position of excretory pore, vulval lips (barely protuberant or highly protuberant) and body size. The more or less marked constriction observed in the pharyngeal corpus-isthmus varied among individuals; these differences were not mentioned in descriptions of other species of the genus. A similar situation occurred with the variability observed in vulval lips. The position of the excretory pore is a very important character for discriminating Thelastoma spp. ( Carreno 2007). In the present work, two positions were observed: at half of the pharyngeal basal bulb (the most frequent situation) and slightly anteriorly, near the isthmus. In the characterization of a population of T. basiri, Singh (1987) found differences from the original description of the species, which the author attributed to the effects of fixation on individuals. However, in females of T. vanwaerebekei n. sp., the two mentioned positions were observed both in live and fixed nematodes, therefore discarding that the differences were due to fixation artifacts.
Body size of T. vanwaerebekei n. sp. females was found to be variable at intra-population level, with individuals that doubled their size. Camino et al. (1997) and Rao (1958) observed a considerable variation in body size in T. rara and T. icemi , respectively. Variability among populations of a single species from different hosts were also found; females of T. longicaudata ( Meyer, 1896) Travassos, 1929 removed from a diplopod ( Julus sp.) were almost twice as big as nematodes obtained from Periplaneta americana L. ( Singh & Malti 2003).
The presence of a seminal receptacle on the female reproductive branch has been mentioned only in a few species of the genus. Thelastoma vanwaerebekei n. sp. is characterized by the presence of a seminal receptacle on both branches. In some species ( T. madecassum , T. tampoketsii van Waerebeke, 1970 , T. toxi , T. icemi , T. retrovulvaris Adamson, 1987 , T. patellae , T. ritteri and T. endoscolium Poinar, 1978 ), illustrations suggest the presence of a seminal receptacle in some of the branches, but this structure was not mentioned in the descriptions. In T. pterygoton , the seminal receptacle is present at the tip of uterine branches; the seminal receptacle is absent on the anterior branch of the reproductive tract in T. dessetae . In T. gipetiti van Waerebeke, 1987 , the anterior seminal receptacle is reduced or absent, and the posterior one is present, suggesting that the posterior seminal receptacle might be due to a precocious development (van Waerebeke 1 987), whereas in T. gueyei , the anterior seminal receptacle is small, sac-like, and the posterior seminal receptacle is reduced or absent ( Koubková et al. 2006). The eggs produced in the anterior branch are not fertilized and would produce haploid males, whereas females would be produced from fertilized eggs (diploid) generated in the posterior branch ( Adamson 1985). This characteristic would be widely distributed in the genus Thelastoma and is assumed to be an adaptation to haplodiplody, which would ensure a given number of males and females in the progeny ( Adamson 1985, 1990).
In vivo, some males of T. vanwaerebekei n. sp. showed irregular protuberances near the anus; similar structures were indicated in the anterior region of males of T. toxi , this being the only species within the genus with records of cuticular protuberances. Furthermore, transverse lines composed of small protuberances were detected ventrally, in the posterior and midbody region. Adults of T. pterygoton frequently exhibited rough areas on their cuticule that were visible under the light microscope; electron micrographs of the affected areas showed numerous microorganisms closely appressed to the cuticular surface ( Poinar 1973). The author suggested that the microorganisms (maybe some type of bacteria or mycoplasma-like body) were capable of dissolving at least part of the nematode’s cuticule and establishing colonies on the surface of the nematode.
The taxonomy of the genus Thelastoma is problematic due to the high number of species that have been inaccurately characterized. Many of these descriptions have been based on scarce specimens or have been poorly illustrated. In addition, there is a lack of adequate diagnostic or taxonomic information on invertebrate hosts, as well as the contradictory lists of valid and invalid species that have been proposed by various authors ( Carreno 2007). Female characters were mostly used for identification ( Baruš & Koubková 2002); however, differentiation was based on males’ characteristics on some occasions ( Farooqui 1970), although they are difficult to find or are unknown for many species. Male thelastomatoids are often difficult to identify due to a lack of informative morphological characters and the sexual dimorphism of the species; as multiple species guilds are common in thelastomatoid infections, matching males to females is difficult, thus impeding the use of male thelastomatoids to discern taxonomic relationships ( Jex et al. 2006). Therefore, in the present work molecular studies considering males and females of T. vanwaerebekei n. sp. were conducted to ensure that both corresponded to the same species.
Accurately identifying individuals of the family Thelastomatidae is fundamental for the study of their systematics, population genetics and ecology ( Jex et al. 2006). Few molecular studies have been performed with species of the genus Thelastoma . There is available information of the 18S SSU rRNA gene only for 4 species: T. vanwaerebekei n. sp., T. icemi ( Chaudhary et al. 2013) , T. krausi ( Nadler et al. 2007) , and T. gueyei ( Koubková et al. 2006) . The highest similarity (98%) was obtained between T. vanwaerebekei n. sp. and T. krausi while T. gueyei (95%) separated from the rest of the species of the genus in the phylogenetic tree. Thelastoma gueyei has high genetic similarity (99%) to Brumptaemilius justini Adamson & Anderson, 1985 , a nematode species from the family Carnoyidae ; this suggests that a revision should be carried out to corroborate the identity of that species.
There are also few partial sequences of the 28S gene corresponding to unidentified Thelastoma species ( Spiridonov & Guzeeva 2009; Blanco et al. 2012), T. bulhoesi ( Ozawa et al. 2016) and a sequence (191 bp) belonging to T. icemi (Singh et al. unpublished). In spite of the scarce molecular data, the rRNA LSU separated T. vanwaerebekei n. sp. from the available Thelastoma spp. (85% genetic similarity), Hammerschmidtiella spp. (74% genetic similarity) and Leidynema spp. (74–77% genetic similarity). These results suggest that LSU seems to be a more useful marker for phylogenetic analyses in Thelastomatidae ; however, additional molecular studies should be performed considering other species to infer the relationships within this family.
Given the taxonomic problems within the genus, it would be important to make a review of all the described species, complemented with scanning electron microscopy. Knowledge on hosts, their distribution, life cycle and feeding mode should also be considered. These aspects are unknown for many species and are necessary for the understanding of the nematode-host relationship.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |