
Tanacetipathes Opresko, 2001
|
publication ID |
https://doi.org/ 10.5281/zenodo.170924 |
|
publication LSID |
lsid:zoobank.org:pub:891DAE3D-595A-4778-AB51-1E69F1015268 |
|
DOI |
https://doi.org/10.5281/zenodo.6266669 |
|
persistent identifier |
https://treatment.plazi.org/id/277F2F3E-7121-FF8A-2642-A0994972FE98 |
|
treatment provided by |
Plazi |
|
scientific name |
Tanacetipathes Opresko, 2001 |
| status |
|
Genus Tanacetipathes Opresko, 2001 View in CoL a
Antipathes Pourtalès View in CoL (1880:116, in part); Brook, 1889:160 ("species incertae sedis"); Opresko, 1972: 959 (in part).
Tanacetipathes Opresko, 2001a:349, 2001b: 358 View in CoL –361.
Diagnosis: Corallum monopodial or branched. Stem and branches with bottlebrush pinnulation to the third or fourth order. Primary pinnules arranged in four to six regular rows and in alternating biserial groups of two or three pinnules each. Secondary pinnules located on lateral sides of primaries and often developed more extensively on abpolypar side. Tertiary pinnules, when present, usually on abpolypar side of secondaries, and sometimes also on polypar side. Quartenary pinnules present in some species.
Type species: Antipathes tanacetum Pourtalès, 1880 (see Opresko, 2001a: 349).
Type material: The type series of T. tanacetum (Pourtales) consists of 12 specimens in the Museum of Comparative Zoology (Harvard University, USA) and two specimens in the National Museum of Natural History (Smithsonian Institution, USA). [It should be noted that four of the MCZ specimens and two of the USNM specimens were identified as being part of Pourtalès type series only after the Opresko’s publication in 1972]. All of these specimens are monopodial, meaning that although they are pinnulate, there are no branches developing from the stem. Based on the length of the primary pinnules and the number and arrangement of the secondary pinnules, there appear to be as many as four different species represented in the type series. Three syntypes have a pinnulation pattern similar to that of T. barbadensis (Brook) with relatively long posterior primary pinnules (2–3 cm) and only a few secondary pinnules near the base of the primaries; three other syntypes resemble T. thamnea (Warner) in having relatively long posterior primaries with secondaries occurring both uniserially and biserially. The remaining eight syntypes have secondary pinnules arranged primarily (but not exclusively) uniserially; four have relatively short, curved primary pinnules (less than about 1.5 cm long) with 2–3 secondaries on the posterior primaries, and the other four specimens have somewhat longer posterior primary pinnules (2 cm or more), with up to 8 secondary pinnules. Pourtales did not designate a holotype for T. tanacetum , nor did he indicate which form he thought was the most “typical” of the species. Because of the similarities of some of the syntypes with other nominal species, it is necessary to select a lectotype from one of the two groups with predominantly uniserial pinnulation. A specimen with short curved primary pinnules was chosen because it has a very distinctive pinnulation pattern that has been previously associated with T. tanacetum ( Opresko, 1972, 2001b).
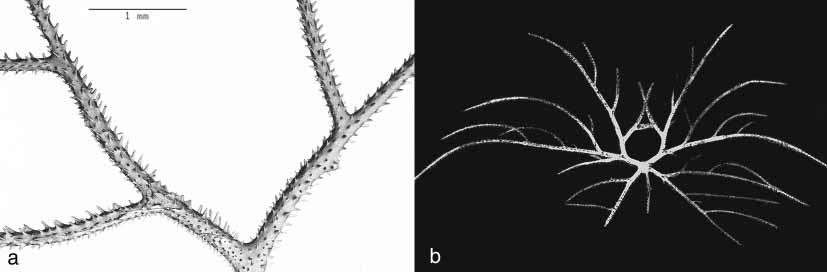
Designation and description of lectotype of T. tanacetum : The specimen selected as the lectotype ( MCZ 57361) is a small colony only 11 cm tall ( Fig. 1 View FIGURE 1 a), with a width across the pinnules of about 2 cm. The colony is not branched. The primary pinnules are arranged biserially mainly in four rows; however in places a few pinnules corresponding to a 5th and 6th row also occur. Within each row the primary pinnules are spaced about 2 mm apart (about six per centimetre). They are also arranged in alternating biserial groups of two or three pinnules, one from each row on each side of the axis ( Fig. 1 View FIGURE 1 b). The posterior primary pinnules are the longest and the anterior primaries (corresponding to the 5th and 6th rows) are the shortest. The posterior primary pinnules are up to 1.2 cm long and about 0.25 mm in diameter near the base (excluding spines); most are broken off at the tip. The middle primary pinnules are generally not more than 7 mm in length and the rarely occurring anterior ones are at most 3 mm. There are one to three secondary pinnules on the posterior primary pinnules; these are usually arranged uniserially on the abpolypar side of the axis ( Fig. 2 View FIGURE 2 a), although occasionally a secondary will also occur on the polyp side of a primary. The secondaries closest to the base of the posterior primaries are up to 6 mm long with a basal diameter of about 0.16 mm. In some cases a basal secondary is longer than the primary on which it occurs. The more distal secondaries are up to 4 mm long and have a diameter of about 0.14 mm. Secondary pinnules (mostly one or two) also occur on the middle primaries where they are usually on the abpolypar side of the axis; however, in places one may occur on the polypar side, in which case it often forms, with the lowermost secondary from the abpolypar side, a subopposite pair. The small anterior pinnules lack subpinnules. Tertiary pinnules, mostly one or two, but sometimes three, are present on many of the secondaries (fig. 2b). Again, the most common condition is for the tertiaries to occur on the abpolypar side, but here and there one may be on the polypar side, or is directed distally or basally relative to the direction of the stem. Rarely, a pair of tertiary pinnules is subopposite. Tertiary pinnules are most often about 3 mm in length or less; however, a few are as long as 5 mm and are longer than the secondary on which they occur. The polypar spines on the primary pinnules are 0.08–0.12 mm tall, as measured from the middle of the base to the apex ( Figs 3 View FIGURE 3 a, b and c). The abpolypar spines are 0.07– 0.09 mm. Some of the spines possess small, flattened, oval to elongated papillae on the middle of their surface ( Fig. 4 View FIGURE 4 a). The polyps are very poorly preserved; however, in places they appear to be 0.6 to 0.8 mm in transverse diameter. A similar polyp size has been recorded for other specimens assigned to this species (see Opresko, 1972), and in the latter specimens the density of the polyps ranged from ten to 13 per centimetre. On the pinnules the polyps are arranged uniserially, and usually occur on the side of the axis with the tallest spines.
Comments: As in other species of the genus Tanacetipathes , the primary diagnostic features of T. tanacetum pertain to the size, number, and arrangement of the subpinnules, particularly in the predominantly uniserial placement of the secondary and tertiary pinnules on the abpolypar side of the lower order pinnules (see Fig. 2 View FIGURE 2 b for a typical arrangement of the pinnules and subpinnules). Other characters that contribute to the definition of the species are: (1) the usual monopodial growth form of the colony, although in some rare cases there may be one or more branches developing from near the base of the stem: (2) the relative shortness of the primary pinnules, with the posterior primaries usually being no more than about 2 cm in length (rarely up to 2.5 cm); and (3) the tendency for both primary and higher order pinnules to be curved back along their course (although this does not occur in all specimens assigned to this species). In large colonies there is usually more extensive subpinnulation (with up to six secondary pinnules on the largest primaries, and sometimes with a quaternary pinnule occurring on the lowermost tertiary. Even so, the basic pinnulation pattern is the same as that occurring in the lectotype. What does vary substantially from specimen to specimen is the size of the spines. As noted previously ( Opresko, 1972), the maximum size of the polypar spines in many specimens, as in the lectotype, fall in the range of 0.10 mm to 0.13 mm, whereas in others it is greater than about 0.20 mm, with few colonies having spines of an intermediate size. This may simply be due to incomplete sampling or it might be indicative of a more significant difference between these two forms. There is no consistent correlation between the size of the colony and the maximum size of the spines, and larger colonies may have either small or large spines (see Opresko, 1972). This suggests the possibility of two separate species; however, until additional biological and ecological and perhaps molecular data are collected on the two forms, it is suggested that the recognized limits of the species include substantial variability in the size of the spines.
Distribution: As the species is defined here, its distribution ranges from the Gulf of Mexico south to Brazil. It has also been reported from Bermuda.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Tanacetipathes Opresko, 2001
| Pérez, C. D., Costa, D. L. & Opresko, D. M. 2005 |
Tanacetipathes
| Opresko 2001: 349 |
Antipathes Pourtalès
| Opresko 1972: 959 |
| Brook 1889: 160 |