
Araeosoma anatirostrum, Anderson, Owen F., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3609.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:046410E8-079F-4FBF-B0EF-361DFCE92E2C |
|
DOI |
https://doi.org/10.5281/zenodo.5622878 |
|
persistent identifier |
https://treatment.plazi.org/id/275FE573-6D7D-FFE1-8C99-FF57FC5D3C16 |
|
treatment provided by |
Plazi |
|
scientific name |
Araeosoma anatirostrum |
| status |
sp. nov. |
Araeosoma anatirostrum View in CoL sp. nov.
Figures 3 View FIGURE 3 F, 4G, 12–15
Holotype —From the seamount Christable (a seamount later closed to trawling) west of the Auckland Islands, to the south of New Zealand: 51° 3.2´S, 164° 36.4´E, 990– 973 m (123 mm TD). Deposited in the NIC, Wellington, NIWA29442. Collected on 14 April 2003 by RV Tangaroa during research into the biodiversity and ecology of seamount fauna (voyage TAN0306), using an epibenthic sled. Stored in 80% ethanol.
Paratypes —Christable seamount: 1 specimen (126 mm TD), 51° 3.2´S, 164° 36.4´E, 990– 973 m, NIWA81373; 1 specimen (63 mm TD), 51° 4.3´S, 164° 36.4´E, 1065– 1030 m, NIWA29443; Macquarie Ridge: 3 specimens (103, 106, 110 mm TD), 50° 15.1´S, 163° 32.1´E, 1033 m, NIWA23708; 2 specimens (112, 113 mm TD), 50° 5.8´S, 163° 28.5´E, 1070–1123 m, NIWA39833. NIWA23708 stored dry, the rest stored in 80% ethanol.
Other material —Chatham Rise (Shipley seamount): 5 specimens (102, 103, 106, 108, 111 mm TD), 41° 48.6´S, 179° 30.9´W, 1251–1290 m, NIWA53314; 3 specimens (99, 103, 110 mm TD), 41° 47.9´S, 179° 30.3´W, 1219–1286 m, NIWA53337; 1 specimen (107 mm TD), 41° 48.1´S, 179° 29.6´W, 1240–1275 m, NIWA25963; 3 specimens (100, 114, 115 mm TD), 41° 48.1´S, 179° 29.6´W, 1240–1275 m, NIWA25969. South Kermadec Ridge (Haungaroa seamount): 1 specimen (112 mm TD), 32° 35.8´S, 179° 36.5´W, 1252– 1175 m, NIWA4663; South Tasman Rise: 2 specimens (114, 117 mm TD), 47° 28.0´S, 148° 55.0´E, 1022 m, NIWA23710; 1 specimen (135 mm TD), 48° 37.5´S, 150° 27.0´E, 913–1148 m, NIWA29445; 1 specimen (123 mm TD), 47° 6.0´S, 148° 42.0´E, 1076–1128 m, NIWA29446. Bounty Plateau: 1 specimen, 47° 17.4´S, 177° 55.3´E, Te Papa EC9361. Tasmania: 1 specimen, 44° 20.5 147 10.6´E, 1593 m, AM J24540 View Materials . NIWA25963, NIWA23710, and one specimen in NIWA25969 stored dry, the rest stored in 80% ethanol.
Etymology —Named anatirostrum , from the Latin anas (duck) and rostrum (beak), after the distinctive duck bill-shaped valves of the tridentate pedicellariae. Used as a noun in apposition.
Diagnosis —Adults small to moderate size, up to 135 mm TD; test and all appendages light brown in colour; relatively narrow membranous spaces between interambulacral plates, wider orally; primary tubercles on every plate in oral adradial interambulacral series; ambitus markedly rounded; spines fragile, frequently lost or broken; hoofs white, short and little flared; two types of tridentate pedicellariae, both with distinctive duck-bill-shaped valves; dactylous pedicellariae not found.
Description —Test of holotype ( Figure 12 View FIGURE 12 ) medium size (test diameter 123 mm), circular to slightly pentagonal, flattened, but ambitus considerably rounded. Test flexible, moderately robust, the spines relatively fragile—many primary spines missing or broken. Colour of ethanol-preserved test pale brown, lighter on the connective tissue between plates, and an intermediate colour on spines and tube-feet—all parts generally of a similar hue, within a shade or two of each other. The colour is slightly variable between individuals, at least in preserved specimens, some being darker than the holotype. Tubercles non-crenulate, primary and larger secondary tubercles perforate. Ratio of interambulacrum to ambulacrum width at the ambitus, about 3:2.
Interambulacral columns with 36–38 plates, about 17 on the oral side and 19–21 aborally. Ambulacral columns with about 60 plates, a few more on the aboral side than the oral side—exact numbers difficult to determine as the sharply angled interambulacral plates cross the very rounded ambitus. Some variability exists between individuals in the number of plates per column: the paratype from the same station as the holotype (NIWA81373) has several more plates in both the interambulacra (43–45) and ambulacra (68–69); specimen NIWA25969 (dry) has 44–46 interambulacral plates and 59 ambulacral plates; and the three specimens in NIWA23708 have 42–46 interambulacral and 62–72 ambulacral plates. Although thick skin partially conceals exact positions of plate boundaries, membranous gaps evident in the median area between successive interambulacral plates. These gaps slightly wider on the aboral side, and relatively narrow compared with other Araeosoma species. This character much clearer in the dried specimens, which show little or no gap between ambulacral plates. Coronal plates typically oblong and widened, where necessary, to accommodate large tubercles.
Oral test plating ( Figure 13 View FIGURE 13 ): A regular series of adradial primary tubercles on every plate in the interambulacra, as in most Araeosoma species, ending at the ambitus. Areoles mostly circular, only a few compressed by or overlapping with adjacent areoles in the series. A second, irregular, series of primary tubercles near the interradial suture on every second plate, sometimes on every third plate or on adjacent plates, from the ambitus to the peristome. These tubercles similar in size to the adradial series adambitally, but becoming relatively smaller adorally. A few smaller primary tubercles also present on a few of the adambital plates between these two series. In some specimens, e.g. NIWA23708 (dry), these form a further series, approximately equidistant between the other two, occurring on every second plate or every plate adorally, the tubercles becoming smaller towards the peristome. These two minor series not always distinct, and variable in tubercle size and placement, but visible to some extent in all individuals examined. In the ambulacra, primary tubercles limited to the interporiferous zone, where there is a single, irregular series in each column, roughly parallel to and near the perradius. The tubercles in this series occur on adjacent plates or on every second, third, or fourth plate, often with a secondary tubercle maintaining the series on plates without a primary tubercle. A series of secondary tubercles lies closely adjacent to this series, squeezed in between it and the inner pores, from the ambitus to about two-thirds down the column to the peristome. This series inconspicuous, occurring on every second or third plate, but continuing onto the aboral side where it is the most prominent series. Other secondary tubercles of similar size lie between the accessory plates and the pore-pair of the main plate on most plates. Smaller secondary tubercles and pedicellariae-bearing granules are scattered over the rest of the coronal plates.
Peristome 29 mm in diameter, bearing only ambulacral plates, 9–10 per column. This number varying slightly between individuals, being 10–11 or 11–12 in some specimens. Peristomial plates only slightly angled and overlapping only marginally with those of the adjacent column; each plate with 3 or 4 tubercles on either side of the central tube-foot, plus one or two granules bearing pedicellariae. Spines curved, slightly expanded distally, and flattened. Gills small, composed of 3 or 4 squat lobes; buccal notches moderately shallow.
Aboral test plating ( Figure 13 View FIGURE 13 ): In the interambulacra, the median series of the oral side continues onto the aboral side as the most prominent series of primary tubercles, situated slightly adradial to the centre of the plate and occurring on adjacent plates or every second plate, to about ¾ up the column to the apical system. A few other large tubercles placed adradial to this series on the outermost few plates, but not forming a distinct series. Interambulacral plate pairs angled at about 90° to each other where they fit around the apical system, but opening further down the column to be about 140–160° at the ambitus, then further so as to be virtually parallel over much of the oral surface. This arrangement typical of all specimens examined, but there exists some variation in degree. For example, the interambulacral plates in the dry specimen in NIWA23708 are all nearly parallel except for those very close to the apical system. The pattern of tuberculation in the aboral ambulacra similar to that of the oral side, but with smaller primary tubercles. The main series follows on from the secondary series of the oral side, as described above, with only one or two other primary tubercles elsewhere on the outer plates. Secondary tubercles occur fairly regularly on either side of the accessory plates on many plates. Smaller secondary tubercles and pedicellariae-bearing granules scattered over remainder of aboral plates, as on the oral surface, but distinctive narrow naked areas aborally—centred on the adradial and interradial sutures.
Accessory plates located more or less towards the adradial end of the primary plate in each compound ambulacral plate, especially orally. Tube foot of the primary plate located near the adradial end, slightly adapical and sometimes with a secondary spine adradial to it.
Apical system monocyclic, 24–25 mm across, the outline pentagonal to star-shaped, exact plate structure difficult to determine due to the covering of connective tissue. Madreporite oval (about 6 x 4 mm), not much raised, indented slightly where encroached by the gonopore, and pierced by 2 or 3 spinelets. In the dried specimens plate G2 is enlarged considerably compared with the other genitals to accommodate the madreporite. Shape of genital plates not determined (but are irregular triangles in dried specimens, pointing down the interradii), a ring of 7–8 spinelets about 3 mm long surrounding the membranous central space into which the gonopores (about 1 mm diameter) open. Genital plates appear divided in some of the dried specimens, especially NIWA25963 and NIWA23710, in which they comprise two to several individual plates. Ocular plates (irregular diamond-shaped in dried specimens) with 5–6 spines; anal plates (several) with a total of about 10 spines. Anus raised slightly into a small cone.
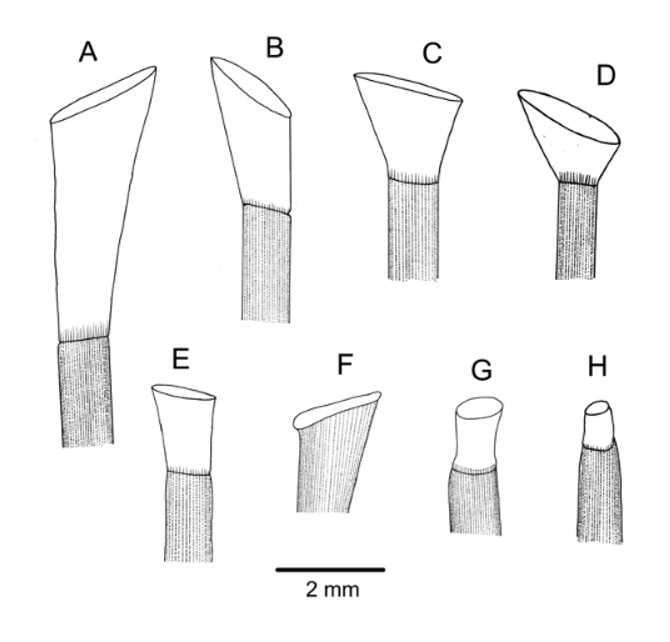
Spines: The spines mostly broken, the longest remaining about 15 mm long (intact spines on other specimens of similar size up to 17 mm long). Primary spines circular and hollow, very delicate; the largest about 0.7 mm diameter, very slightly curved and with about 30 fine, longitudinal striations. These striations simple club-shaped wedges in cross-section in the aboral spines examined, but wedges with lateral extensions in the oral spines; the spine walls quite thin—a ring or two of perforations bordering the inner wall ( Figure 3 View FIGURE 3 F). Several hoofs found, on both intact and broken spines. These hoofs white and relatively short (maximum length of about 2 mm) and only slightly, if at all, flared ( Figure 4 View FIGURE 4 G).
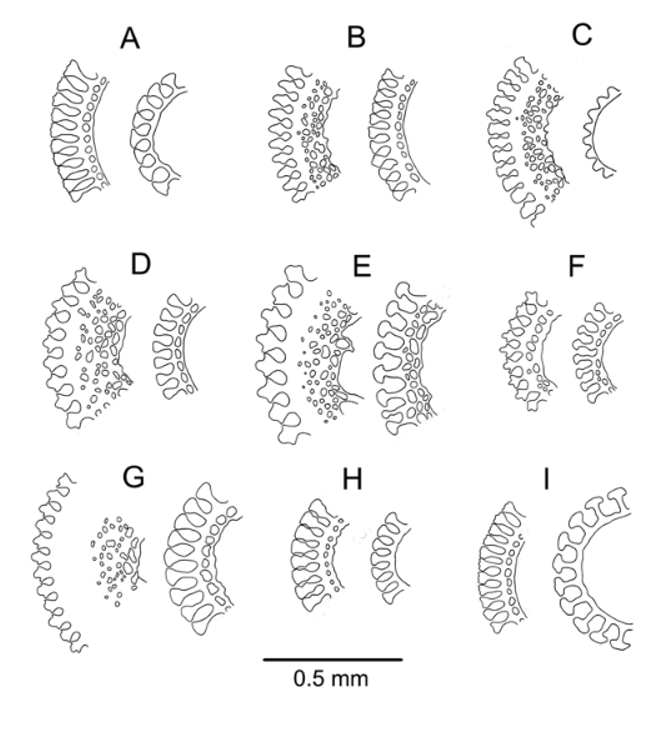
Pedicellariae ( Figure 14 View FIGURE 14 ): Two types of tridentate pedicellariae present: The most common type by far is a rostrate form, with distinctly duck-bill shaped valves, narrowest in the middle and widening towards the evenly rounded, very finely serrated tip. These occur in a variety of sizes from about 0.5 mm to 1.5 mm valve length, the blades in contact for most of their length, the neck quite short (up to about ¼ head length), and the stalk long. Blades more or less filled with meshwork and typically strengthened by a longitudinal septum. The other tridentate is a larger (about 2 mm blade length) involute type, not at all common, in which the valves touch only at the extremities of the widened tip and wide gaps separate the blades. Blades narrowed in the middle and widened distally, but this widened section finely serrated below the point of contact at the tip and angled so as to be well designed for clasping relatively large objects. One example was 4-valved, but this was perhaps an anomaly as no others were found on the holotype or other specimens. A few partially developed tridentate pedicellariae were also found, sharply pointed with triangular valves lacking a strengthened border, on very short stalks, similar to those observed in specimens of other species (e.g. A. thetidis ). Triphyllous pedicellariae unremarkable, with a typically long neck and blade involuted so as to be enclosed to a varying degree. These are usually about 0.5–0.6 mm long (although one example was found of about half this length) the blade tip about twice as wide as the base. The holotype and other type material were searched for dactylous pedicellariae, but none found; these pedicellariae tend to be rare and difficult to find in other Araeosoma species, so are not necessarily absent from A. anatirostrum , and may eventually be found in other specimens.
Sphaeridia elongated, club-shaped (about 1.0 mm long), transparent, located adjacent (slightly perradial and adoral) to the inner tube foot in each ambulacral plate.
Size range —The median test diameter of the 24 specimens measured was 110 mm, and the largest specimen was 135 mm TD.
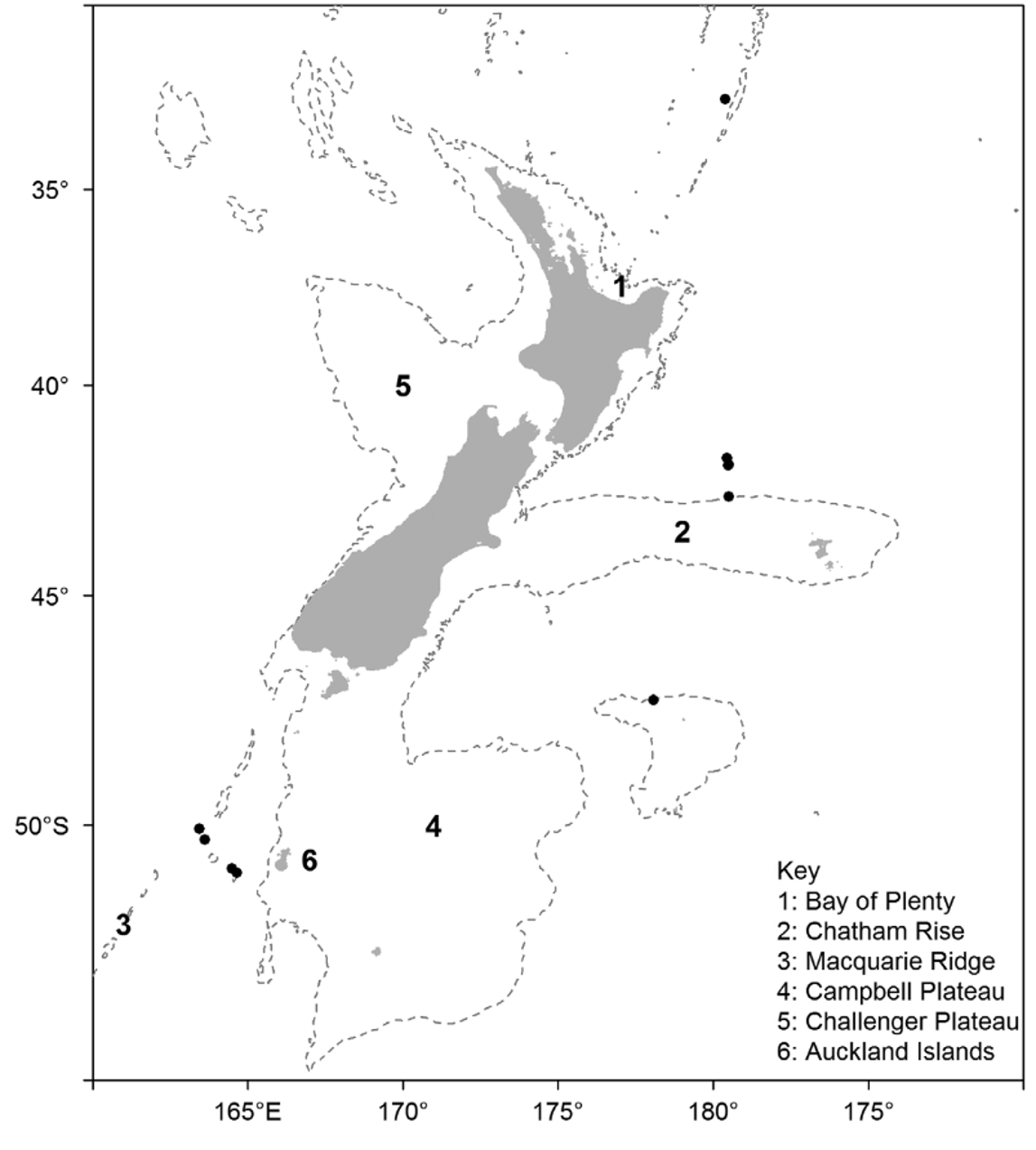
Occurrence —There have been 14 confirmed records of A. anatirostrum since the first specimen was taken in 1997, representing 28 individuals.
These records are spread widely over the New Zealand region as well as south-eastern Australia ( Figure 15 View FIGURE 15 ); most are in the vicinity of seamounts or other undersea features associated with bottom trawling for deepsea fish such as orange roughy, alfonsino, and oreos. The New Zealand specimens (and some of the South Tasman Rise specimens) were mostly collected by government fisheries observers working on board vessels in these fisheries, or by scientists on board RV Tangaroa studying these fisheries or the ecology of the seamounts; one record from south of Tasmania was taken by the CSIRO research vessel RV Southern Surveyor. Nearly half of the specimens came from three research trawls on an isolated seamount (Shipley seamount) north of the Chatham Rise, and a third of the specimens came from four stations further south, on Christable seamount and adjacent regions of the Macquarie Ridge. The species is not limited to sub-Antarctic waters however, with one record from the southern Kermadec Ridge, to the north of New Zealand. This is the deepest of all the New Zealand Araeosoma species, with captures mostly from the extreme of the range of current trawl fisheries, and it may be that this species is relatively common at these less frequently sampled depths.
The conservative depth range for the species is 990–1251 m with a potential range of 913–1593 m.
Remarks —The general appearance of the test of A. anatirostrum is of the form where the aboral interambulacral plates are strongly angled to each other, fitting around the elongated genital plates, as also seen conspicuously in A. bakeri sp. nov. The very rounded ambitus is also a distinctive feature of the species. The tuberculation patterns are typical for Araeosoma .
The species most similar to A. anatirostrum is probably A. leptaleum (Agassiz & Clark, 1909) . The regular tridentate pedicellariae of A. anatirostrum look similar to the figure of such a pedicellaria in Mortensen (1935), taken from the holotype of A. leptaleum , but none match the description of Agassiz & Clark (1909) which includes forms similar to those of A. thetidis and Sperosoma giganteum . The tuberculation patterns in A. anatirostrum are similar to those of A. leptaleum (see Agassiz & Clark 1909, Plate 76) but differ in that the inner columns of tubercles in the oral interambulacra are less distinct in A. leptaleum , the conspicuous naked median area in the aboral interambulacra in A. leptaleum is not present in A. anatirostrum , and the zig-zag pattern of alternating tubercles in the aboral interambulacral plates of A. leptaleum is not apparent in A. anatirostrum . The characteristic feature of A. leptaleum , as noted by Mortensen (1935) and by Pawson & Ahearn (2001) from specimens collected from near the Galapagos Islands, is the shape of the hoof, which is distinctly tapered rather than flaring—a form unique in echinothurids. In A. anatirostrum the hoof is narrow, but slightly flaring. Having examined Pawson & Ahearns specimens, and also Agassiz & Clark (1909) figures, it is also clear that the membranous gaps between plates, especially on the aboral side, are much wider in A. leptaleum .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Echinothuriinae |
|
Genus |