
Araeosoma leppienae, Anderson, Owen F., 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3609.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:046410E8-079F-4FBF-B0EF-361DFCE92E2C |
|
DOI |
https://doi.org/10.5281/zenodo.5622882 |
|
persistent identifier |
https://treatment.plazi.org/id/275FE573-6D64-FFE9-8C99-FA3DFE4A3F2D |
|
treatment provided by |
Plazi |
|
scientific name |
Araeosoma leppienae |
| status |
sp. nov. |
Araeosoma leppienae View in CoL sp. nov.
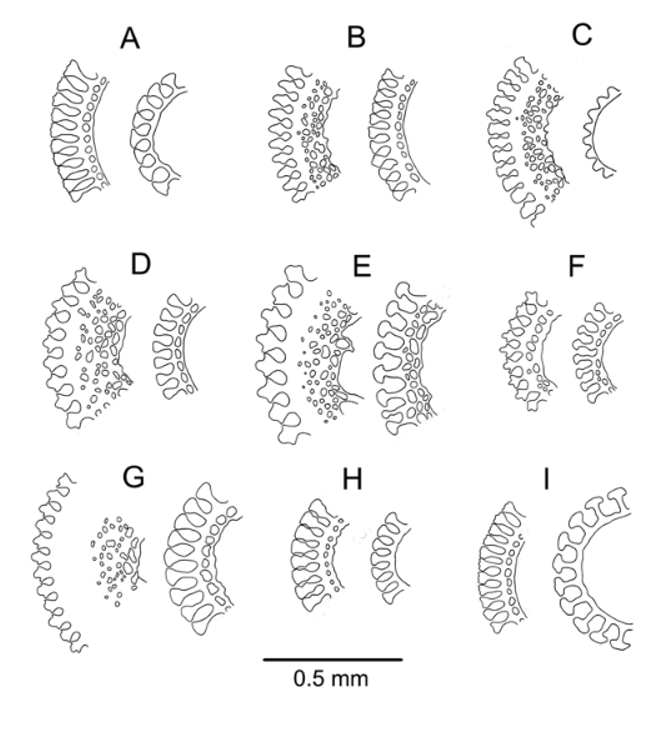
Figures 3 View FIGURE 3 I, 20–22
Holotype —From Christable seamount, west of the Auckland Islands, south of New Zealand: 51° 3.9´S, 164° 35.3´E, 968–973 m ( 118 mm TD), NIWA23724. Deposited in the NIC, Wellington. Caught on 14 April 2003 by RV Tangaroa during research into the ecology of seamounts. Stored in 80% ethanol.
Paratypes —None.
Etymology —Named after Bridget Leppien, used as a noun in the genitive case.
Diagnosis —Test and spines brown, darker in poriferous zone and paler between columns creating a radial pattern of alternating light and dark stripes; large tridentate pedicellariae with broad, coarsely serrated valves; small tridentate pedicellariae narrow, valves not serrated; involute tridentate pedicellariae lacking; dactylous pedicellariae with six valves; primary tubercles on every plate in the oral adradial interambulacral series.
Description —Test of holotype ( Figure 20 View FIGURE 20 ) moderate size ( 118 mm TD), circular, flattened, rounded at ambitus. Colour of test brown, darkest in the pore zones partly due to retention of colour in the tube-feet and their protection of the surrounding skin during capture, lighter in the interambulacra and interporiferous zones where test plates are more net-worn. This produces a pattern of radial stripes on both surfaces—a feature which may not be as apparent in the live animal. Primary spines all broken and mostly lost, therefore details of the hoofs unknown. Primary tubercles perforate and non-crenulate. Ratio of interambulacrum to ambulacrum width at the ambitus, 3:2.
Interambulacral columns have 35–36 plates, 15 on the oral side and 20–21 aborally; ambulacral columns have 51–57 plates, 20–23 orally and 31–34 aborally. Plate boundaries obscured by thick membranous connective tissue but wide gaps clearly evident between interambulacral plates on both surfaces. Tubercles less prominent at inner end of aboral plates due to partial covering by connective tissue, accentuating broad naked areas interradially and perradially. Areoles of primary spines large, creating the highest point of the plate, but mostly not overlapping when adjacent on neighbouring plates. In each primary plate, at least those away from the poles, the adoral accessory plate is adjacent to the adapical accessory plate of the neighbouring plate in the usual way for echinothurioids (see Mortensen 1935, Mooi et al. 2004), approximately in mid-plate position. The pore-pair of the main plate lies slightly separate from these pairs, adapical to them and closer to the outer end, those in the adambital plates regularly separated from the abambulacral margin by a conspicuous secondary tubercle. Closer to the poles the pore-pairs are cramped into the available space among these smaller primary plates. Aboral plates slightly shorter than oral plates, in accordance with their greater number; aboral interambulacral plates only slightly angled to each other.
Oral test plating ( Figure 21 View FIGURE 21 ): Adradial series of primary tubercles in the interambulacra of the usual arrangement, with a tubercle on every plate from the ambitus to the peristome. An inner series is located close to the interradius, with a tubercle on every second plate near the ambitus but more frequently on every plate as the tubercles diminish in size and move toward a mid-plate position adorally. A few smaller primary tubercles also found in the distal part of the oral surface, in between the two main series. An irregular series of primary tubercles runs down the perradial margin in the ambulacra, with tubercles of variable size occurring on adjacent plates or every second or third plate. One column in the zone cleaned for examination has noticeably more large tubercles than the other—the large tubercles in one column corresponding with plates lacking a primary tubercle in the other. A large secondary tubercle lies adradial to these primary tubercles on most plates, but not in any regular position, and one to several other secondary tubercles form a rough line along each plate.
Peristome about 27 mm in diameter, bearing two overlapping columns of 8–9 slightly angled (ambulacral) plates in each zone with an extra small plate amongst the distal-most plates. Each plate bears a central tube-foot flanked by a row of 4–5 tubercles spanning the width of the plate. Buccal notches shallow; gills relatively large, lobate.
Aboral test plating ( Figure 21 View FIGURE 21 ): No continuation of the oral adradial series of primary tubercles onto the interambulacra of the aboral side, this series ending abruptly at the ambitus. Instead, a distinctive series of primary tubercles beginning slightly outside mid-plate at the ambitus and becoming more adambulacral adapically, quite regularly on every second plate, ends about two-thirds of the way to the apical system. A few other primary tubercles and larger secondary tubercles present near the interradius, approximately continuing the oral inner series, over the distal half of the aboral surface. A series of smaller primary tubercles in the ambulacra, located usually on every second or third plate, runs adjacent to the inner tube-feet in similar fashion to the oral series but placed a little further from the perradius and comprising more equal-sized tubercles. An additional primary tubercle also found near the adambulacral end of a few of the outer plates.
Apical system compact, about 18 mm across, skin obscuring plate boundaries. The whole system evenly covered with about 50 small spines and pedicellariae (mostly triphyllous). Genital pores small, opening within a membranous space; genital plates not much extended into the interradii, therefore adjacent interambulacral plates not much angled to fit around them. Madreporite distinct, slightly raised, subtriangular. Anus central.
Spines cylindrical, hollow, fragile; the longest remaining section about 10 mm long, 1 mm in diameter, and smooth with about 35 longitudinal striations. These striations simple club-shaped wedges in cross-section in the oral spines examined, but the wedges more mallet-shaped in the aboral spines. Spine walls thin—a row of perforations in the oral spine wall but strengthening meshwork completely lacking in the aboral spines sectioned ( Figure 3 View FIGURE 3 I). Colour of both primary and secondary spines uniformly brown, the colour fading in bleach. Milled ring not prominent, only very slightly oblique.
Pedicellariae ( Figure 22 View FIGURE 22 ): Tridentate (large and small), triphyllous, and dactylous forms present. Tridentate: Large rostrate form (about 1.0 mm long), valves wide with a very broad, coarsely serrated tip and only slightly narrowing towards the base, blades filled with strengthening meshwork. The neck 1–2 times head length, the stalk 3–4 times head length; small rostrate form (about 0.7–0.8 mm long) with narrow, straight-sided, and unserrated blades with a rounded tip, neck about equal to head length, stalk several times head length. No involute tridentate pedicellariae present. Triphyllous: typical echinothurioid form about 0.5–0.6 mm long on a long neck and stalk. The basal part completely enclosed along much of the blade in the examples seen. Dactylous: of the highly developed form typical of the genus, about 2.5 mm long, with six valves. These are relatively common near the ambitus on the aboral side.
Sphaeridia of the usual form, i.e., elongate spherical (about 0.7 mm long by 0.2 mm wide), located slightly perradial and adoral to the inner tube foot of the ambulacral plates.
Remarks —Although there is only a single specimen recorded for this species, it is sufficiently distinct from the other Araeosoma species known from the region ( A. migratum , A. anatirostrum , and A. bidentatum ) to be certain that it is not simply an unusual form of one of them or a hybrid of two of them. Interestingly, however, A. anatirostrum is also found at Christable seamount (usually in slightly deeper water), its holotype being caught on the same day as that of A. leppienae . The large tridentate and dactylous pedicellariae of A. leppienae are of a form unique among the New Zealand Araeosoma species. The dactylous pedicellariae are particularly unusual in that no other species of Araeosoma is reported to have more than five valves.
The forms of pedicellariae found in A. leppienae are probably most similar to those of the Indonesian A. parviungulatum (another species known from a single specimen), which also has no involute form. Both species have one large and one small rostrate tridentate form, as well as triphyllous and dactylous forms. The two species differ in that the large tridentate pedicellariae of A. parviungulatum are more pointed than that of A. leppienae , the small tridentate pedicellariae of A. parviungulatum have a distinct waist which is not present in A. leppienae (see plate LXXVII in Mortensen 1935), and the dactyous pedicellariae are five-valved in A. parviungulatum and sixvalved in A. leppienae . Details of colour, tuberculation patterns (especially aborally), and other test characters also distinguish the two species.
The type locality, Christable seamount, is adjacent to the Macquarie Ridge, a major undersea ridge feature running south from New Zealand to about 60˚ S, which would likely provide a considerable amount of similar habitat. Relatively little sampling using methods which could capture individuals of this species has taken place in the area, and so it is probable that A. leppienae occurs in suitable habitats along this ridge and other sub-Antarctic ridges and seamounts.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Echinothuriinae |
|
Genus |