
Araeosoma tertii, Anderson, Owen F., 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3609.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:046410E8-079F-4FBF-B0EF-361DFCE92E2C |
|
DOI |
https://doi.org/10.5281/zenodo.5622880 |
|
persistent identifier |
https://treatment.plazi.org/id/F2ACE0B3-2E44-41D2-9943-26FA3637003B |
|
taxon LSID |
lsid:zoobank.org:act:F2ACE0B3-2E44-41D2-9943-26FA3637003B |
|
treatment provided by |
Plazi |
|
scientific name |
Araeosoma tertii |
| status |
sp. nov. |
Araeosoma tertii View in CoL sp. nov.
Figures 3 View FIGURE 3 D, 4E, 16–19
Holotype —From Tuatoru Knoll in the Bay of Plenty, northeast New Zealand: 37° 28.2' S, 177° 13.2' E, 200– 175 m ( 165 mm TD), NIWA6615. Deposited in the NIC, Wellington. Caught on 12 November 2004 by RV Tangaroa during research into the ecology of seamount fauna. Stored in 80% ethanol.
Paratypes —All from Mahina Knoll in the Bay of Plenty, collected on the same research survey as the holotype: One specimen ( 157 mm TD), 37° 21.3' S, 177° 6.0' E, 260–280 m, NIWA6600; one specimen ( 141 mm TD), 37° 28.3´S, 177° 13.5´E, 218– 200 m, NIWA6618; one specimen ( 170 mm TD), 37° 28.3´S, 177° 12.3´E, 295– 200 m, NIWA6614; one specimen ( 135 mm TD), 37° 28.3´S, 177° 12.9´E, 180– 177 m, NIWA6616. Stored in 80% ethanol.
Other material —One specimen ( 170 mm TD), from the West Norfolk Ridge, northwest New Zealand, 33° 46.0' S, 167° 13.0' E, 313 m, NIWA45138, stored in 80% ethanol; one specimen ( 91 mm TD), from north of Norfolk Island, 28° 42.3´S, 167° 56.7´E, 475 m, NIWA29453, stored dry; two specimens ( 140, 150 mm TD), from the east coast of northern North Island, 35° 25.2´S, 175° 0.4´E, 270 m, NIWA23718, stored dry.
Etymology —Named tertii , the Latin translation of the Māori word Tuatoru (meaning “third”) after the type locality Tuatoru Knoll. Used as an adjective in the genitive case.
Diagnosis —Adults large, up to 170 mm TD; test and appendages deep red; few large primary tubercles aborally; prominent membranous spaces between interambulacral plates; primary tubercles on every plate in the oral adradial interambulacral series; large tridentate pedicellariae with broad, coarsely serrated valves; small tridentate pedicellariae narrow but broadening gradually towards tip; dactylous pedicellariae not found.
Description —Test of holotype ( Figure 16 View FIGURE 16 ) large (test diameter 165 mm), circular, flattened, ambitus rounded. Colour of test a deep, dark red, owing to the pigment in the skin and plates; pale pink membranous connective tissue between the plates. The white hoofs and sucking discs of the oral tube-feet contrast strongly with the red, producing a striking appearance. Primary tubercles perforate and non-crenulate. Ratio of interambulacrum to ambulacrum width at the ambitus, 5:3. Paratypes similar size to holotype ( 140–174 mm TD).
Interambulacral columns have 45–46 plates; 19 on the oral side and 26–27 aborally. Ambulacral columns have 67–68 plates; 29–30 on the oral side and 38 aborally. As usual in this genus exact plate boundaries obscured by thick membranous connective tissue. This tissue tends to exaggerate gaps between adjacent interambulacral plates, especially aborally, but such gaps clearly evident on both surfaces (dried specimens not available to accurately examine this feature). This membranous tissue also produces broad naked areas between columns of plates, especially interradially. Areoles of primary spines large, those in adjacent plates within the same series mostly touching or overlapping.
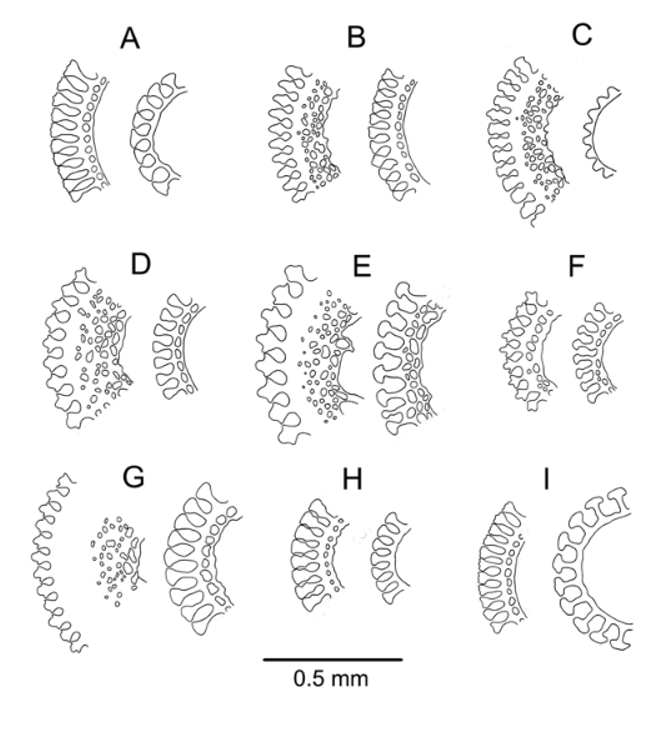
Aboral test plating ( Figure 17 View FIGURE 17 ): On the outermost 1–4 interambulacral plates only, there is a primary tubercle on the adradial margin, often skipping a plate—a continuation of the oral series in this position. In addition, a much smaller primary tubercle is found on most plates—in no repeated regular pattern but mostly on the adradial half of the plates—from the ambitus to the apical system. These plates otherwise scattered with a few secondary and miliary tubercles, and granules bearing pedicellariae. In the ambulacra, primary tubercles form an irregular series close to the perradial end, occurring on every second or third plate throughout the column. No other large tubercles elsewhere on these plates, but a few secondary and miliary spines, and granules, scattered about. Tube-feet of primary plates located at adapical, adambulacral corner adapically, but noticeably further away from adambulacral plate margin in outer third of aboral surface. Tube-feet of accessory plates lie adjacent in an adoral position near plate centres. Aboral plates short and wide compared with oral plates, in accordance with their greater number; interambulacral plates not much angled to each other, except those near the apical system.
Apical system monocyclic, about 23 mm across; plate boundaries obscured by thick covering of skin. Genital pores small, opening in membranous space within the disaggregated genital plates. Genital plates not extending much into interradii, requiring only the adapical-most interambulacral plates to be angled around them. Ocular plates with 2–3 small tubercles; ocular pore located at outer edge of plate. Madreporite distinct, slightly raised, quite circular (about 7.5 mm in diameter) and split clearly into 2 large and 3–4 very small sections. Anal tube prominent—much expanded distally to form a broad cone shape. Remainder of apical system covered with numerous small plates, each with 1–4 small tubercles of varying size—this more obvious in the dried specimen, in which it can also be seen that these plates form a rough ring around the central anus.
Oral test plating ( Figure 17 View FIGURE 17 ): A regular series of adradial primary tubercles on every plate in the interambulacra—the usual arrangement for Araeosoma —increasing only gradually in size outward from the peristome and not continuing past the ambitus. An inner series of similar sized tubercles occurs on adjacent or every second plate, located near the perradius adambitally, but becoming mid-plate adorally. In other specimens, e.g., NIWA45138, this series differs in that the tubercles occur on more of the plates, sometimes nearly all. An additional primary tubercle lies adjacent to the perradial margin on one or two plates in each column (more frequently in a few other specimens, e.g., on every second or third plate in NIWA6600) and also on one or two plates adjacent to the primary adradial series. Several smaller secondary and miliary spines also scattered over the plates. A series of primary tubercles in the ambulacra, quite regularly on every second or third plate, lies near the perradius. A series of secondary tubercles discernable near outer end of plates, within the pore zone, on adjacent plates or every second plate. As on the aboral surface, outer tube-feet adjacent to plate edge on central plates but distinctly further away adambitally. Small spines cover remainder of plates, as elsewhere on the test.
Peristome about 38 mm in diameter and bearing only ambulacral plates, each with a central tube-foot surrounded by a row of 2–6 large tubercles and several small tubercles which span the width of the plate. Two overlapping columns of 14–15 slightly angled plates in each zone, several of which are paired in the distal part. Buccal notches distinct; gills moderate size, palmate.
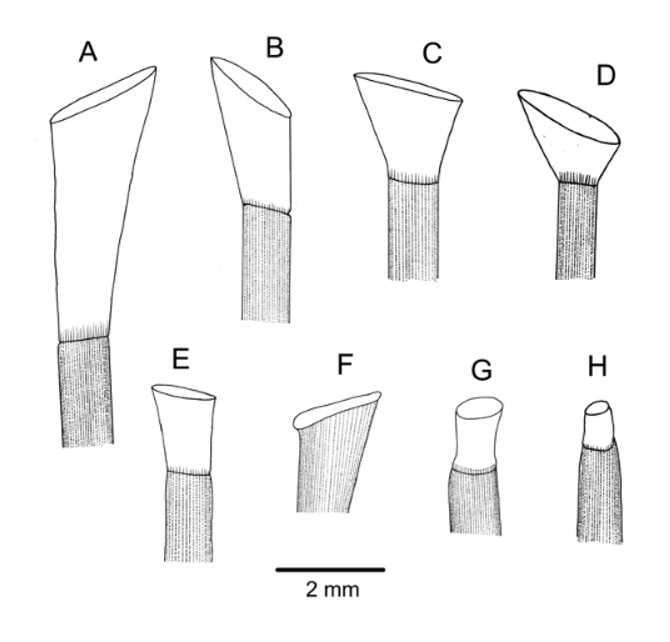
Spines cylindrical, hollow, milled ring distinct but not prominent, usually slightly oblique. Primary spines smooth, the largest with about 40 fine, longitudinal striations. These striations simple club-shaped wedges in crosssection in the aboral spines examined, but wedges with lateral extensions in the oral spines. Oral spine wall with a relatively thick layer of meshwork; aboral spine wall thin, enclosing a single row of perforations ( Figure 3 View FIGURE 3 D). Colour of all spines a uniform red, the colour being in the skin and readily bleached out. Oral spines curved, the longest about 28 mm with a diameter of about 1.0 mm. Hoofs white, moderately long (up to about 2.5 mm), slightly flared, and slightly flattened distally ( Figure 4 View FIGURE 4 E).
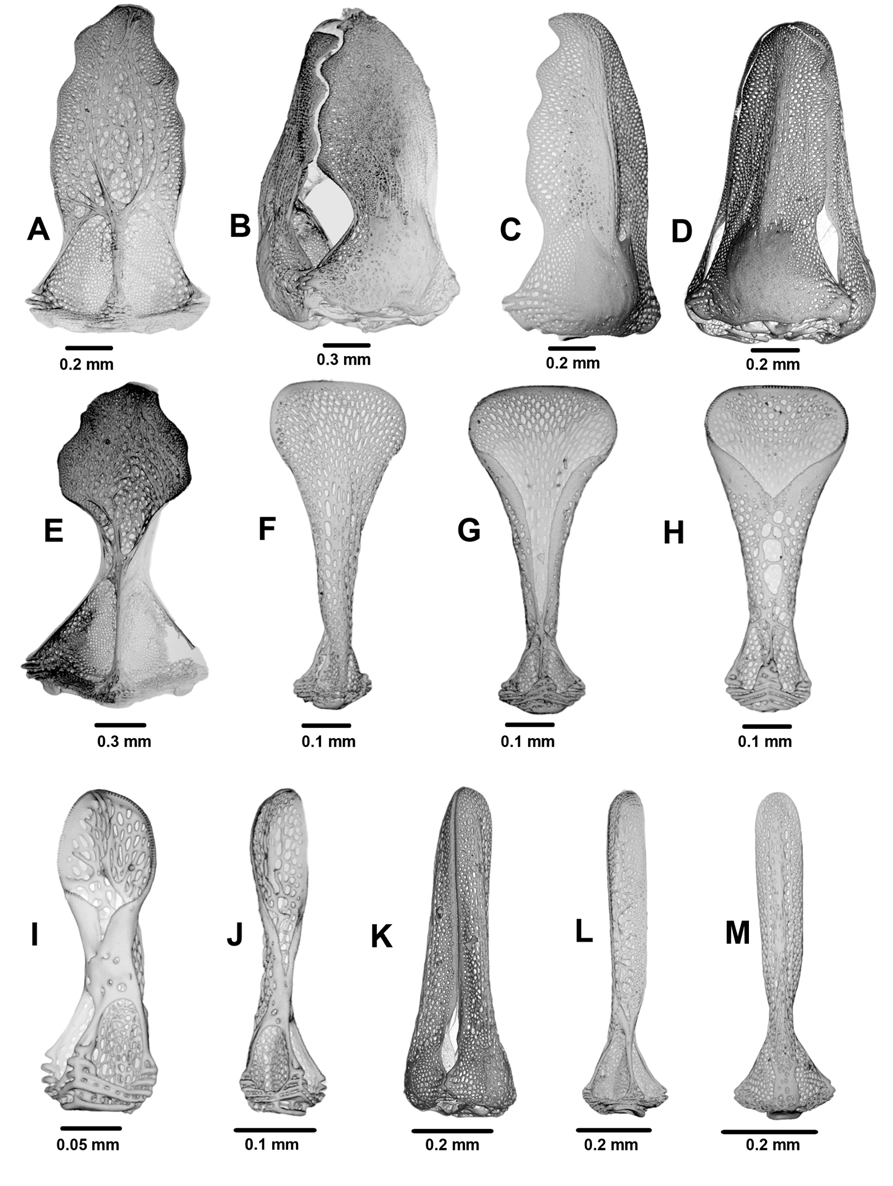
Pedicellariae of two types ( Figure 18 View FIGURE 18 ): tridentate (a rich variety of large and small) and triphyllous. Large tridentates ( 1.2–2.4 mm) usually quite common, ranging from short, wide involute forms with wide gaps between blades in the closed head to longer, rostrate forms of variable width and with only narrow gaps between blades, and intermediate forms. Blades coarsely serrated, some only at the tip and others over the entire outer part, and usually partly filled with strengthening meshwork. Small tridentates with narrow, un-serrated and finely pointed valves, outer side of blade with a distinct longitudinal ridge. Triphyllous pedicellariae the typical echinothurioid form, about 0.7 mm long on a long neck and stalk. Extent to which basal part of the valve completely enclosed highly variable. One or two smaller specimens (about 0.25 mm) with a short blade also found, but these may be semi- or mal-formed. Dactylous pedicellariae not found.
Sphaeridia elongate spherical (about 0.5 mm long by 0.2–0.3 mm wide), located adjacent to the inner tube foot in each ambulacral plate, slightly perradial and adoral to it. None found on the aboral surface.
Size range —Median test diameter of the 9 specimens measured 146 mm, the largest specimen 170 mm TD.
Occurrence —There are just eight records of A. tertii (representing nine individuals), all from northern New Zealand (mainly the Bay of Plenty) and Norfolk Island ( Figure 19 View FIGURE 19 ). The species is strongly associated with rocky substrates, with five of the records from seamounts surveyed on a single research voyage by RV Tangaroa . Whether A. tertii is restricted to this region or whether the known records define the southern part of a more tropical distribution can only be answered by further sampling north of New Zealand in appropriate depths along the major ridge features and among the islands of the southwest Pacific.
The conservative depth range for the species is 180–260 m, with a potential range of 175– 475 m.
Remarks —There are several other red echinothuriinids which are superficially quite similar to A. tertii . The species most similar is perhaps A. splendens , an Indonesian species known from a single specimen taken from depth of 300 m. This similarity is most apparent in the colour of the test and spines and in the general forms of the pedicellariae present. The two species otherwise differ in aspects of the test plating and tuberculation, and in the finer details of the pedicellariae. Although the tridentate pedicellariae are broadly similar between the two species the distinct longitudinal keels on the blades in A. splendens are not present in A. tertii . Also, in A. tertii , the larger tridentate pedicellariae have a more expanded blade and the smaller tridentate pedicellariae usually have a distinct waist—the blade gradually broadening towards the tip rather than being straight sided. The ambulacral tuberculation is fairly similar in the two species, with irregular series on both surfaces, but in A. tertii these series continue further towards the apical system. Further differences are seen in the adradial series in the aboral interambulacra, which is not restricted to the distal part in A. tertii as it is in A. splendens , and in the much smaller contrast in the number of ambulacral and interambulacral plates between the oral and aboral surfaces in A. splendens .
The pedicellariae of A. tertii , particularly the involute tridentate forms, are also quite similar to those of another Indonesian species, A. tessellatum . In that species too there is a great variation in size and shape in the pedicellariae (as figured by Mortensen 1935, Plate LXXXI). Details of the test plating and colouration are otherwise quite different in these two species.
In colouration A. tertii is also similar to Calveriosoma gracile , but the pattern of tuberculation in that species is much more regular, particularly orally, and its distinctive involute tridentate pedicellariae have a much finer blade tip than any of those found on A. tertii . There is also a striking similarity between A. tertii and Hapalosoma amynina sp. nov., but this is limited mainly to colour and to macroscopic features of the oral surface, and the species are readily separated by examining the tuberculation of the aboral interambulacra.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Echinothuriinae |
|
Genus |