
Araeosoma alternatum Mortensen, 1934
|
publication ID |
https://doi.org/10.11646/zootaxa.3609.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:046410E8-079F-4FBF-B0EF-361DFCE92E2C |
|
DOI |
https://doi.org/10.5281/zenodo.5622890 |
|
persistent identifier |
https://treatment.plazi.org/id/275FE573-6D50-FFD4-8C99-FB95FCC23F60 |
|
treatment provided by |
Plazi |
|
scientific name |
Araeosoma alternatum Mortensen, 1934 |
| status |
|
Araeosoma alternatum Mortensen, 1934 View in CoL
Figures 3 View FIGURE 3 E, 4C, 29–31
Material examined —West Norfolk Ridge (Hiroshima seamount): 4 specimens ( 175, 112, 192 mm TD), 34° 22.6´S, 168° 25.1´E, 376–380 m, MV F168905. East coast Northland: 2 specimens (150, 124, mm TD), 34° 57.6´S, 175° 10.8´E, 581– 553 m, NIWA3519. Bay of Plenty: Tumokemoke seamount, 1 specimen ( 170 mm TD), 37° 28.1´S, 177° 7.0´E, 230–322 m, NIWA46084 (dry); 1 specimen, 37° 42.0´S, 176° 45.0´E, Te Papa EC763. East Chatham Rise: Diamond Head C, 3 specimens ( 172, 139, 130 mm TD), 44° 8.8´S, 174° 41.4´W, 440–600 m, NIWA53579; Diamond Head B, 2 specimens ( 155, 144 mm TD), 44° 9.0´S, 174° 45.4´W, 519–609 m, NIWA54042. Puysegur Bank: 1 specimen, 46° 40.0´S, 165° 42.0´E, Te Papa EC6607 (dry). Auckland Islands: 1 specimen ( 194 mm TD), 49° 1.0´S, 166° 32.9´E, 620–627 m, NIWA62822 (dry); 1 specimen ( 180 mm TD), 49° 7.3´S, 166° 41.8´E, 571–593 m, NIWA46089; 2 specimens ( 208, 208 mm TD), 49° 17.1´S, 166° 45.7´E, 618–622 m, NIWA65167; 2 specimens ( 230, 182 mm TD), 49° 28.2´S, 166° 37.2´E, 537– 531 m, NIWA61623 & NIWA66710; 4 specimens ( 215, 226, 181, 215 mm TD), 50° 58.0´S, 165° 45.0´E, 549 m, NIWA23728, NIWA29429, & NIWA29436; 1 specimen, 49° 55.3´S, 165° 59.7´E, Te Papa EC3983 (dry). Macquarie Ridge (Seamount 5): 1 specimen ( 159 mm TD), 51° 3.7´S, 161° 58.7´E, 385–498 m, NIWA39933. Tasmania: 1 specimen ( 205 mm TD), 44° 4.0´S, 146° 24.1´E, 470–524 m, MV F168937; 28 specimens, 44° 9.1´S, 147° 7.7´E, 800–950 m, AM J24562 View Materials ; 2 specimens, 44° 12.6´S, 147° 2.6´E, 1078– 593 m, AM J24542 View Materials . Madagascar: 3 specimens ( 118, 120, 130 mm TD), 12° 51.0´S, 48° 8.0´E, 558 m, MNHN IE-2007-228. Unless stated, stored in 80% ethanol.
Size range —The median test diameter of the 27 specimens measured was 173 mm, and the largest specimen was 230 mm TD.
Remarks —Before this review, the only previously recorded specimen of A. alternatum was one collected by the German Deepsea Expedition of 1898–99 from off the southern coast of Somalia. In the report on the echinoids from this expedition (Döderlein 1906) the single specimen was mistakenly identified as A. coriaceum . The specimen appears not to have been sighted by any other later taxonomists but, due to Döderlein’s detailed description and figures, Mortensen (1934) was able to identify this error and recognized it as a new species, which he named A. alternatum . The type specimen was originally held in the Berlin Museum but is no longer there (or in the Munich Museum where much of Döderlein’s collection is housed) and it seems likely that it did not survive the second World War (A. Ziegler, MCZ Harvard, pers. comm.).
In 2010, three specimens of an unidentified echinothurioid collected from north of Madagascar were loaned to NIWA from the Paris Museum (MNHN). These specimens (from the same region of the Indian Ocean as the type location of A. alternatum ) matched Döderlein’s description of A. alternatum and also were identical to a number of specimens of a previously undetermined species in the New Zealand and Australian collections examined.
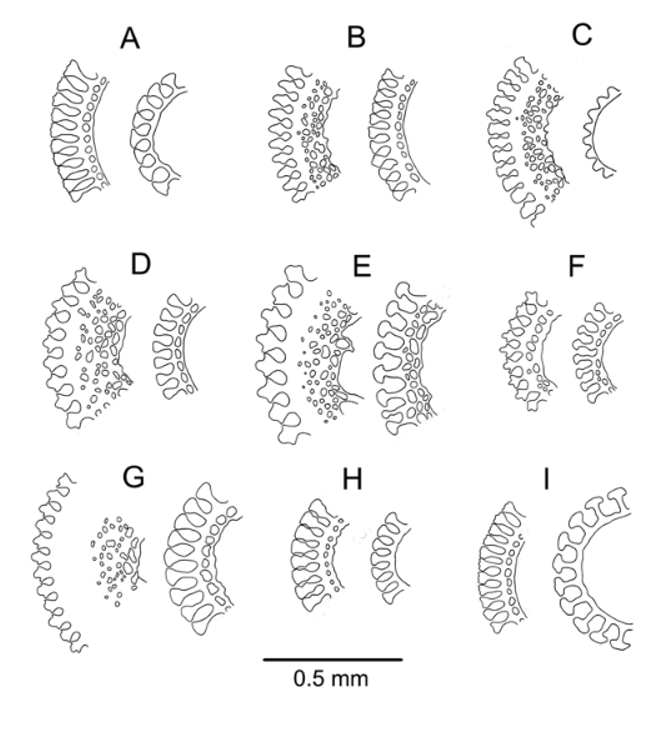
Döderlein’s figures of the pedicellariae of A. alternatum match the pedicellariae of the New Zealand and Australian specimens, and also those seen on the specimens from Madagascar. The large, involute tridentate pedicellariae were relatively common on the specimens examined and very distinctive ( Figure 31 View FIGURE 31 ). The smaller, rostrate tridentate pedicellariae occur in a range of sizes, the broad serrations on the blades diminishing in size with progressively smaller specimens. The unusual and prominent tooth on a distal extension of the apophysis figured by Döderlein was also present in some specimens. The particularly elongate triphyllous pedicellariae are characteristic of this species, and were readily found on the specimens examined. On one specimen from near the Auckland Islands (NIWA66710) a few ophicephalous pedicellariae were found. This is remarkable, as this type of pedicellariae was previously unknown in the genus Araeosoma . This form appears rare within the species too, although they are very small and not all specimens could be thoroughly checked. They resemble more the form seen in Hapalosoma pellucidum rather than those known from other echinothurioids ( Hygrosoma , Sperosoma , and Tromikosoma ) as they have a rounded rather than triangular blade, although in A. alternatum the blade has only a slight medial constriction and is strengthened internally by the extension of the apophysis to the distal end of the blade. That this species should not be reassigned Hapalosoma is fairly certain, however. The large involute pedicellariae of A. alternatum are unknown in Hapalosoma and the tuberculation patterns are more similar to other Araeosoma species. But one of the principal differences between the two genera, the form of the dactylous pedicellariae, cannot be gauged as this type of pedicellaria has yet to be found on A. alternatum .
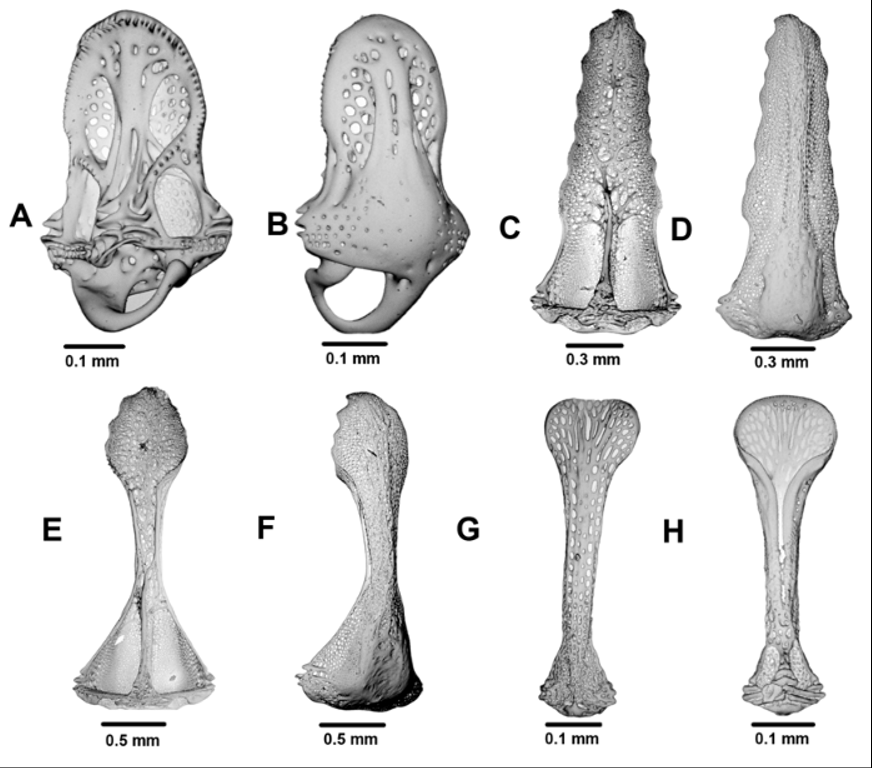
Döderlein was unable to describe the apical system in any detail due to the thick covering of skin, but in two of the dried New Zealand specimens (NIWA3519 & NIWA29429) some details could be made out. These show a pentagonal to star-shaped outline (about 18 mm across in a specimen of 122 mm TD), a monocyclic plate arrangement, and triangular genital plates that extend slightly into the interambulacra. The gonopore opens into an oval or sometimes almost round membranous space, enclosed or nearly enclosed by the undivided genital plate. Plate G2 is enlarged 2–3 times the size of the other genital plates to accommodate the madreporite, which is raised and entirely covers the plate. The anal opening is surrounded by an inner band of about 40 small plates and an outer band of 35–40 larger plates, most with a number of small tubercles or pedicellaria-bearing granules.
This species is one of the largest of the Araeosoma species, the test of the holotype is recorded as 190 mm TD and many of the other specimens examined are a similar size or larger. This species is also distinctive for its very stiff, leathery test; pink or light purple colouration; and the tuberculation of the oral interambulacra which, as in A. thetidis , has a tubercle on every second plate rather than every plate in the adradial series. As explained above, these two species are very similar (several of the A. alternatum specimens examined were originally identified as A. thetidis ) and stand apart somewhat from other Araeosoma species.
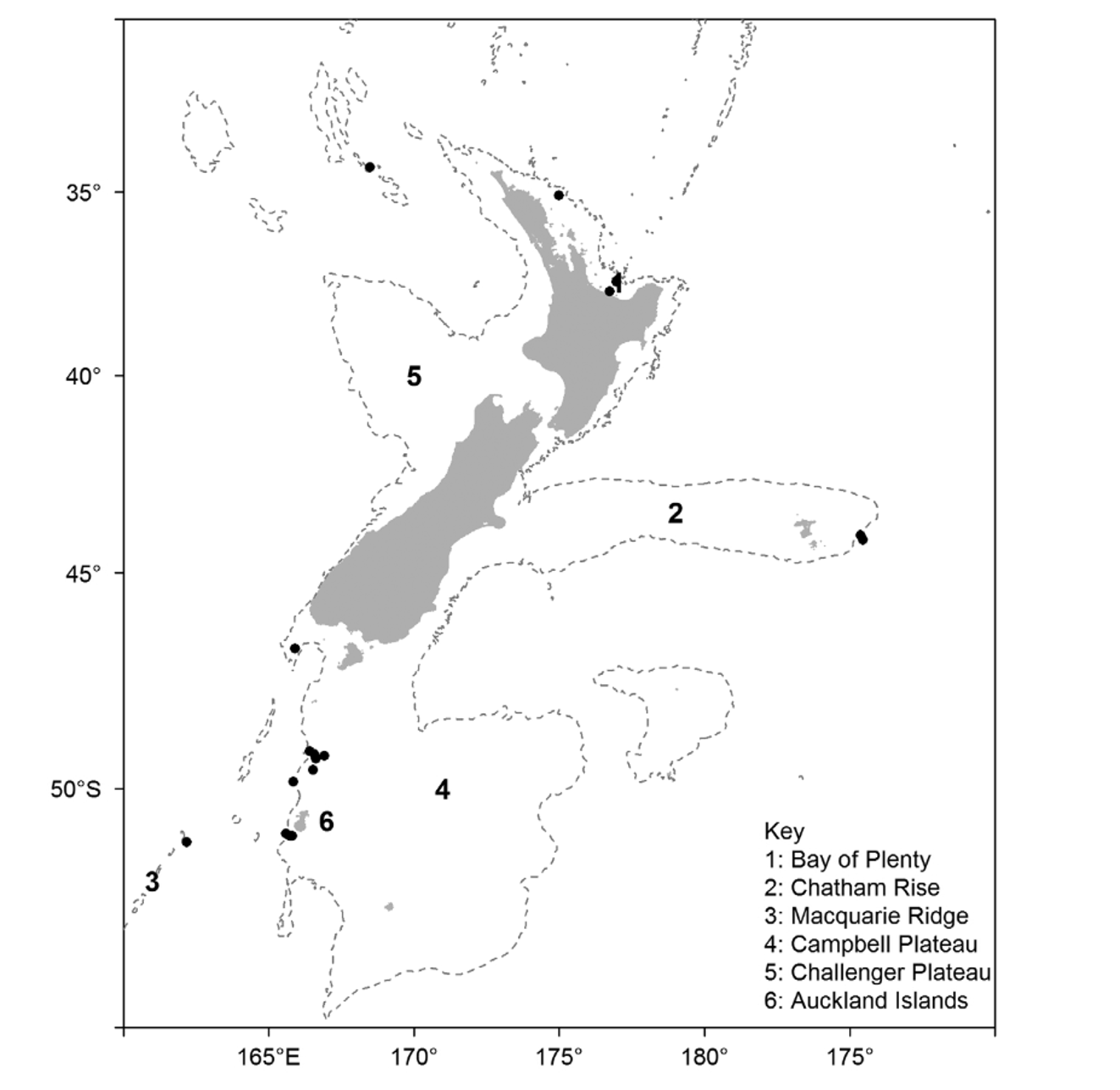
Occurrence —There are 17 confirmed records of A. alternatum from New Zealand and southeast Australia, the first taken from the Bay of Plenty in 1961, representing 59 individuals ( Figure 30 View FIGURE 30 ). In the western Indian Ocean, the species is known from off the Somalia coast (the type locality) and from north of Madagascar.
The Australasian records cover a wide geographical range, from the West Norfolk Ridge in the north to the Macquarie Ridge in the south, and from Tasmania in the west to beyond the Chatham Islands in the east. There are large gaps in the distribution; in particular there are no records from the New Zealand shelf between the Bay of Plenty and the bottom of South Island. It seems likely that the species does occur in this region and the lack of records may be due to a combination of their rarity and the lack of research sampling in the area within the depth range of the species. But by far the largest gap in the distribution of the species is between the western Indian Ocean and Tasmania, a distance of over 11 0 0 0 km. This apparently discontinuous distribution is more difficult to explain, but perhaps the species will ultimately be found to exist further west in Australia and in other parts of the Indian Ocean.
The conservative depth range for the species within Australasia is 380–800 m, with a potential range of 376–1078 m. The depth recorded for the holotype was somewhat deeper ( 1289 m) but given that only a single depth was recorded it is possible that it was living shallower than this. The other Indian Ocean specimens were from well within the depth range found in Australasia.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Echinothuriinae |
|
Genus |