
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/10.5281/zenodo.3942851 |
|
DOI |
https://doi.org/10.5281/zenodo.5503883 |
|
persistent identifier |
https://treatment.plazi.org/id/1C7087B9-6E3C-FFED-FF71-FD11AA15F577 |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
The great theropod Tyrannosaurus rex is usually depicted in an active, bipedal pose, perhaps in pursuit of prey or facing off an opponent. Some artists, e.g., Lawrence Lambe (1917), Gregory S. Paul (1988), John Sibbick ( Norman 1991, p. 72), and Michael Skrepnick ( Currie et al. 2004), have provided views of these animals in other, less active postures, including lying prone or squatting. Presumably the animal would rest with a substantial portion of its body mass supported by the prominent pubic boot. Trace fossils of small crouching theropods show both tarsal and pubic-ischiatic impressions (e.g., Gierliriski et al. 2005). In the great theropods, descending from a standing pose to a rest position was presumably a straightforward matter of squatting, a process considerably less involved than the complex sequencing of folding movements that some modern large quadrupeds, such as camelids ( Gauthier-Pilters and Daag 1981) and bovids, use to lower their mass to the ground. Tyrannosaurus rex might simply have settled vertically in one continuous flexion movement involving the hip, knee, and ankles.
It is in rising from a prone or squatting rest position that some concern for the mechanics of the tvrannosaurid frame might present itself. How could the center of mass (COM) be controlled so that the animal was stable while rising? Was there sufficient mechanical advantage in the major extensor muscles to provide a direct ascent that retraces the trajectory followed in descending to the ground? Were the forelimbs useful in stabilizing the body and in providing thrust during the initial stages of the ascent? To address some of these questions, a fully articulated digital model of Tyrannosaurus rex was created where limb movements are delimited by anatomically based estimates of achievable range of motion, and the position of the instantaneous COM of the animal can be visualized in order to judge balance and stability.
Extant bipeds that might serve as analogs for studying the sitting and standing movements of T. rex , include members of the Macropodoidea, notably the large red kangaroo ( Macropus rufus) and a variety of birds, particularly the large ratites such as the emu (Dromaius novaehollandiae) and the ostrich ( Struthio camelus). As animal mass increases, muscular strategies cannot be expected to scale indefinitely ( Alexander 1989); the effortless rise of a small passerine from rest to a bipedal stance might require multiple, more deliberate stages of limb extension in a biped of several orders greater weight. The biomechanical principles governing the choice of strategy, particularly as regards scaling with body mass, are not well understood. Motion studies have concentrated on capturing relatively steady-state locomotion (e.g., Muybridge 1899: Jenkins etal. 1988), not the transient body movements associated with sitting or standing.
To examine the potential movements that take the animal from standing to sitting, and vice versa, it is important to begin with an estimation of the typical stand and sit postures. Movements that smoothly transition between these extremes can then be proposed and analyzed. In their analysis, it is important to understand how the COM translates during the movement. Longitudinal (caudal-cranial) pitching movements in particular would produce instability that would have to be corrected at risk of injury to the great theropod. It is also important to examine range of motion issues throughout the sit-stand movements and the mechanical leverage of large muscle groups for providing the necessary movements.
Proposals have been offered for how T. rex could sit down on its pubic boot, then rise by first using the forearms as props to help anchor the front of the body while the rear legs were straightened. The upper body would then be tilted back to regain an upright standing posture ( Newman 1970). This idea is but one of the potential uses proposed for the forelimbs ( Osborn 1906; Horner and Lessem 1993; Carpenter and Smith 2001; Carpenter 2002).
In the following, an articulated, 3-dimensional digital reconstruction is used to explore alternative hypotheses regarding the sit-stand movements of this dinosaur. The process of descending and then ascending is amenable to quantitative modeling, taking into consideration the distribution of mass in the animal and the flexibility of those joints involved in the movements, particularly the ankle, knee, and hip within the hind limb, and the potential role of the forelimbs in the process of rising. QuickTime video showing the action is available in the supplemental CD-ROM.
Creating an Articulated Digital Model
DinoMorph software ( Stevens 2002) provides a framework with which to create and pose a digital model of Tyrannosaurus rex . The software can accept 3-dimensional data representing bone morphology (e.g., from computed tomographic [CT] scan or hand digitization), as well as more schematic and simplified representations. In this study, the Tyrannosaurus rex specimen BHI 3033 (Stan) at the Black Hills Museum of Natural History was used as the source for the digital model. The articulation of the appendicular skeleton and the morphology of the pelvic and pectoral girdles were of particular importance, so they were specifically for this study ( Fig. 11.1 View Figure 11.1 ). Digitization data of the head was provided from an earlier CT scan made by Virtual Surfaces Inc. and the Black Hills Institute. The remainder of the axial skeleton was modeled schematically, with centra, neural spines, lateral processes, chevrons, and ribs in a dimensionally accurate but simplified form ( Fig. 11.2 View Figure 11.1 ).
The next step was to estimate the relative placement of each bone within the overall skeletal framework. Along the presacral axial skeleton, the intervertebral separations and overall curvature were determined from measurements and photographs in lateral view. Likewise, the rib cage was formed by painstakingly adjusting each digitally represented dorsal rib to match the curvature, dimensions, and placement of its counterpart in reference photographs that were underlaid within DinoMorph as background images ( Fig. 11.3 View Figure 11.1 ). To refine the 3-dimensional skeletal model, the trunk was successively viewed in anterior, dorsal, and lateral orientations, and for each view, the curvature and placement of the ribs were adjusted so that the digital ribs superimposed precisely over their photographic counterparts. The pelvic girdles, complete with furcula, were then placed on the rib cage as they are mounted on Stan ( Larson and Rigby 2005).
Next, those DinoMorph parameters governing the position and orientation of all appendicular joints were adjusted to create a neutral standing pose, the starting point for this study. Then, for the major appendicular joints important to this study, a range of motion was determined on the basis of an estimate of the thickness and extent of the intervening cartilage in modern avians and direct manipulation of the casts (Kenneth Carpenter and Yoshio Ito, personal communication June 2005). Direct manipulation assisted in determining, for example, the axis of rotation of the femur head within the acetabulum, and in the forelimb the orientation of the fully extended forelimb with respect to the pectoral girdles.
In analyzing potential sit-stand strategies of a theropod dinosaur weighing several metric tonnes, it is important to track the trajectory' undertaken bv the COM during hypothesized movement. The COM is computed in DinoMorph bv assigning both a volume and a density to each discrete segment of the skeleton, such as each interval of the axial skeleton associated with an individual vertebra. Fitted conical and elliptical cylinders are used as a first-order approximation to the body cross-sectional area, governed by adjustable parameters ( Fig. 11.4 View Figure 11.1 ). The density (i.e., specific gravity) associated with each segment was adjusted to roughly reflect cranial and axial pneumaticity, air sacs, and lungs. The COM was computed by summating the gravitational moments associated with each segment throughout the skeleton. By assigning densities of 0.8-1.0 to presacral regions and 1.0 to segments of the appendicular skeleton and caudal vertebral series, the overall COM was located just anterior to the pubic shaft (see Fig. 11.4B View Figure 11.1 ), consistent with estimates by Henderson (1999) and Hutchinson and Garcia (2002). Small but potentially significant shifts in the instantaneous COM during movements could be detected visually as the movement unfolded. For this study, a lithe reconstruction of the cross sections of soft tissue associated with each segment of the skeleton was chosen to corresponding to recent computations by one of us (P. L.). The resulting overall mass for BHI 3033 (Stan) was estimated as ~4400 kg., or about 80% of the mass estimated for the more robust specimen FMNH PR2081 (Sue) by using the same techniques (Stevens et al., in preparation). Although it was possible to estimate as little as 3800 kg for the same skeletal structure by progressively reducing body bulk, particularly in the pelvic region, for this study, the position of the COM was of greater importance than the magnitude.
Reconstructing a Sitting Movement
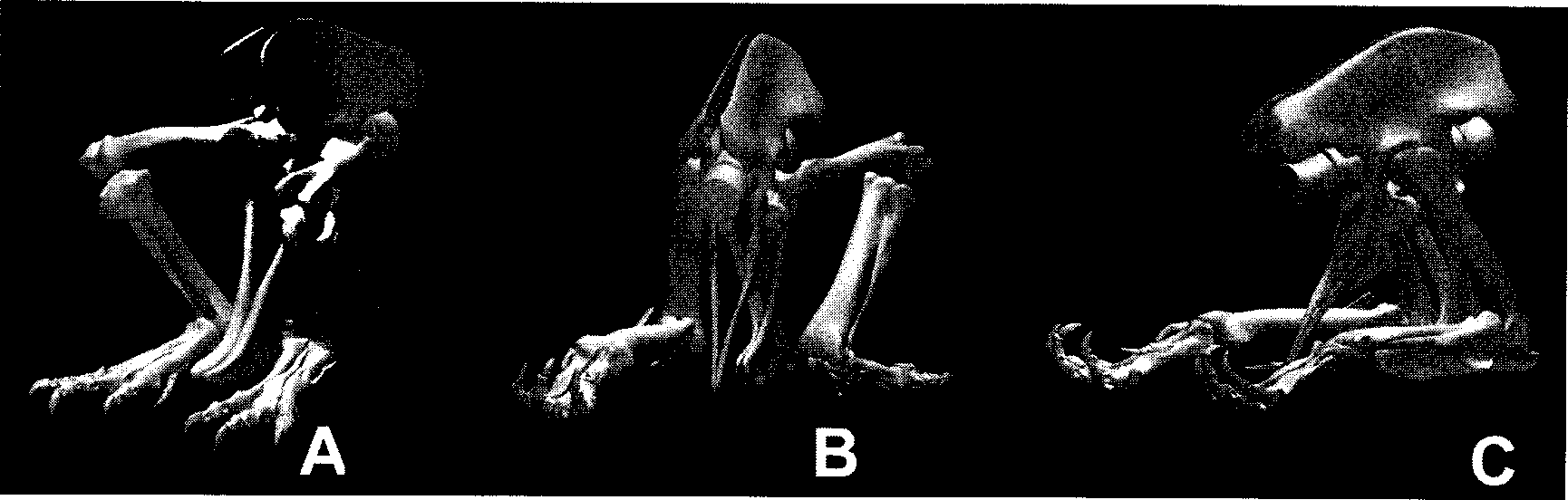
It is reasonable to assume that Tyrannosaurus rex , and other theropods with distally expanded pubic boots, lowered itself until the majority of its mass bore down on the pubis. Upon ground contact, the orientation of pelvic girdle would have shifted slightly so that the elongate ventral surface of the pubis laid generally parallel to the horizontal ground. The elongate pubic shaft of T. rex places the ventral surface of the pubic boot just below the knee, permitting simultaneous ground contact at the knees and along the pubic boot ( Figs. 11.5 View Figure 11.1 - 6 View Figure 11.6 ). The pubis thus likely provided a stable means to offload the great majority of the animal’s weight, limiting pressure on the respiratory system, and to permit repositioning of the hind limbs without requiring a shifting of weight. The limit of hip flexion (femoral protraction) is difficult to estimate because it was governed by soft tissues, but it likely was sufficient to permit achieving the protraction shown in Figure 11.5A View Figure 11.1 so that the tarsus could lie flat on the ground. The limits of knee and ankle flexion are more obvious in the osteology. It is noteworthy that in a full squat ( Fig. 11.5 A View Figure 11.1 ), which brings the pubic boot in contact with the ground, the knee and ankle are nearly fully flexed.
Tyrannosaurus rex , while resting on the pubis, could freely retract one or both femora ( Figs. 11.5 and 11.6 View Figure 11.1 ) and hence vary the stretch on the M. caudofemoralis. Optimal mechanical advantage occurs when the femur is roughly vertical (i.e., associated with the thrust phase in locomotion). Although the femoral position providing greatest muscle moment would correspond to roughly that in Figure 11.5B View Figure 11.1 , the placement of the pes in Figure 11.5 A View Figure 11.1 would appear better suited for elevating bipedally because the hind feet are then un der the COM. If T. rex were not to slowly rise vertically into a stationary standing position, but instead accelerate diagonally from the squat in Figure 11.5 A View Figure 11.1 , then, provided the hind limbs direct the ground reaction force diagonally through the COM, no net pitching moment would be created as the animal rose. Much as a sprinter begins a race accelerating and rising gradually out of the blocks, it is not inconceivable that T. rex could have accelerated diagonally upward from sitting into forward locomotion. Although it is more likely an option for light young tyrannosaurids, it remains a matter of quantitative modeling to estimate whether that was achievable by an adult.
If the femoral retractor muscles were not in an advantageous state for lifting Tyrannosaurus rex vertically1 out of a squat into a balanced standing position, what were the alternatives? One suggestion (Phillip Manning, personal communication June 2005) borrows from modern analogues. In ratites in particular, the M. gastrocnemius comes into play: the Achilles tendon stores energy when in a state of stretch, which is trapped when the animal’s weight bears down on the tarsus while sitting. By leaning forward onto the its knees, the tendon is released, and the hind limb receives a passive boost. Whether recovery of stored mechanical energy would scale to be of significant value in helping boost Tyrannosaurus rex from sitting to standing would require quantitative study. One further concern, beyond the matter of scaling to be effective on a 4000-kg animal, is whether the stored energy would dissipate during the period of rest as the Achilles tendon would stretch.
Another approach is to enlist the forelimbs, as suggested by Newman (1970), as a potential use for these appendages. When sitting, they are close to the ground and are brought into contact by a slight tipping of the pelvis about the prepubis ( Fig. 11.8 View Figure 11.1 ). They could have been instrumental in rising back into a standing position. As shown in Figure 11.9 View Figure 11.1 , although the forelimb range of motion is curiously limited ( Carpenter and Smith 2001), when each forelimb is extended laterally, with elbow straight and manus extended as well, the arrangement resembles a jack stand (or a pair of bicycle kick stands).
The stout forelimbs, fully extended and acting as struts anchored into the ground by strong manual unguals (which are also well oriented for this anchoring task), are well placed for stabilizing the anterior of this giant theropod in preparation for rising. As the animal shifts its weight, ground reaction forces would have been directed nearly perpendicularly into the cup shape of the glenoid fossa; the compressive load would then distribute along the scapulocoracoid over a span of ribs. If the stout forelimbs were indeed involved in stabilizing the body, it is noteworthy that the ground reaction forces would have communicated directly to the vicinity of the acromion process of the coracoid, and therefore place the furcula, which is directly aligned with this force vector, under significant bending stress. As noted by others ( Larson 2001; Larson and Rigby 2005; Lipkin and Carpenter this volume; Rothschild and Molnar this volume), the furcula is frequently found with evidence of healed stress fractures and breaks.
The sprint start discussed earlier would have been assisted bv braced and stabilizing the anterior portion of the body by holding the forelimbs strutlike. Indeed, the resemblance of the initial pose to that of a human sprinter is striking ( Fig. 11.10 View Figure 11.1 ).
Alternatively, with the forelimbs serving to anchor the animal, the posterior musculature of the hind limbs could come into play more gradually to elevate the COM even though it was located ahead of the hind feet. With the animal’s weight positioned fractionally between anchored forelimbs and extending hind limbs, the COM could be elevated with the additional mechanical advantage of a second-class lever. The animal would remain in a stable quadrupedal stance during this initial stage of elevation, and progressively; as the femora and knee come out of the deep crouch, the mechanical advantage of the large femoral retractors and knee extensors would have increased. If not intending a sprint start, but merely wishing to regain a stand- ing posture, the great theropod would likely have (1) tipped forward slightly, pivoting about the prepubis, until (2) the forelimbs were in ground contact and helping to anchor the giant, then (3) it would have first raised its rump, much as large herbivores do today, then (4) either step into forward movement or ascended symmetrically into a standing posture.
Unlike quadrupeds such as bovids, the disparity between forelimb and hind limb length in Tyrannosaurus rex limited the extent to which it could ascend rump first while maintaining a purchase on the ground with the forelimbs. Before achieving full extension of the hind limb, the animal would have had to break contact between its forelimbs and the ground, and either take a step with one hind limb in order to regain its balance, or remain symmetrically posed on 2 hind limbs, and by means of momentum, body movements, and strength, bring the COM back between the hind feet. Perhaps the furcula injuries reflect mishaps that occurred while attempting to regain its balance, particularly when lame as a result of other injuries, or they may reflect the amount of stress imposed on the shoulder girdle during these maneuvers. In the event of a misstep or other failure to achieve balance between the hindfeet, 4 or more metric tonnes falling on the forelimbs could have precipitated such fractures.
Conclusion
The great bulk of an adult Tyrannosaurus rex was capable of being gracefully lowered until it settled its weight on the elongate pubic boot, freeing the animal to adjust its legs much as a sports spectator would use a portable onelegged stool. When it came to rising again to a bipedal stance, the options, particularly for a small tyrannosaurid, would be a sprint start with or without assistance from the forelimbs, or a more gradual elevation using the hind limbs during which the forelimbs played an essential role. The latter was energetically more efficient and might have been preferable for the adult. The forelimbs were literally pivotal in this operation, and mishaps might have resulted in transmission of enormous compressive forces on the pectoral girdles and the delicate furcula that spanned the acromion processes. Although it was perhaps ungainly for the tyrant king to rise rump first, its ascent was likely more elegant than that of modern bovids rising from repose.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |