
Belgopeltula belgica ( Vincx & Gourbault, 1992 ) Holovachov, Oleksandr, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.383 |
|
publication LSID |
lsid:zoobank.org:pub:8D18E64D-6E01-43CC-9A97-BD5C4D2160E1 |
|
DOI |
https://doi.org/10.5281/zenodo.3853261 |
|
persistent identifier |
https://treatment.plazi.org/id/1A10CE2E-C036-FFDD-E386-FAFDFB7495B6 |
|
treatment provided by |
Carolina |
|
scientific name |
Belgopeltula belgica ( Vincx & Gourbault, 1992 ) |
| status |
gen. et comb. nov. |
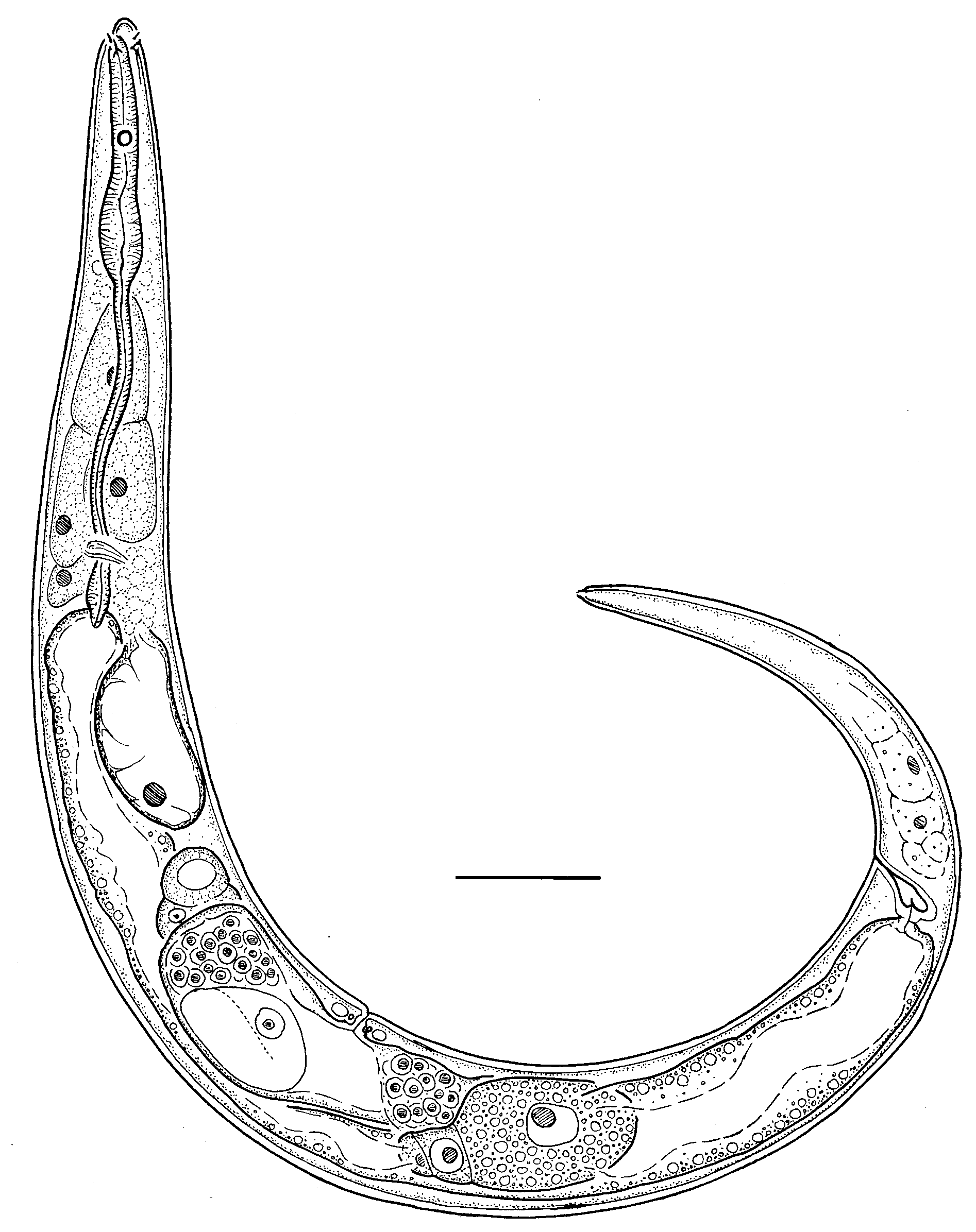
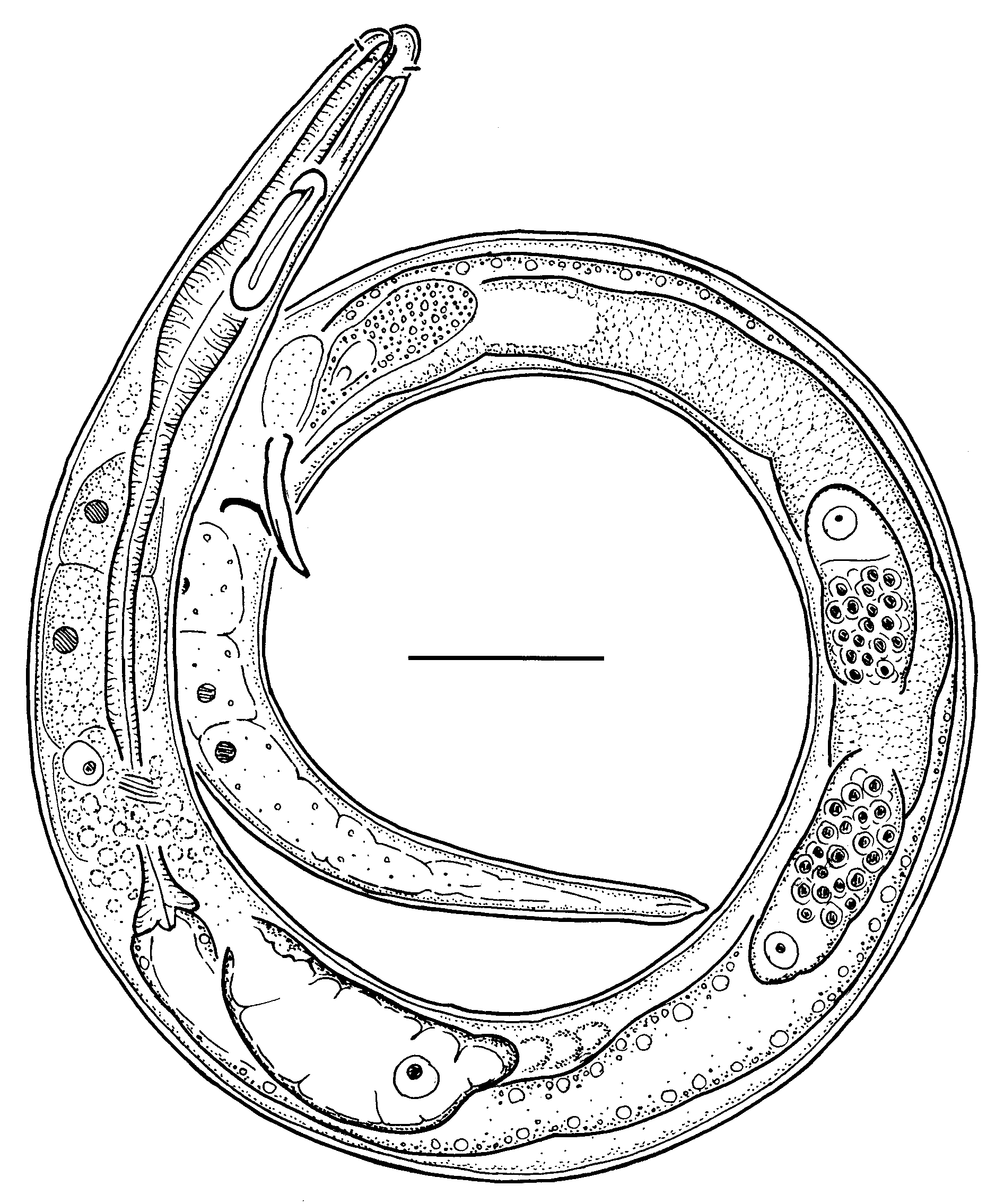
Belgopeltula belgica ( Vincx & Gourbault, 1992) View in CoL gen. et comb. nov. Figs 1–2 View Fig View Fig ; Table 1 View Table 1
Diagnosis (based on all known populations)
Belgopeltula belgica gen. et comb. nov. is characterised by a body of 0.33–0.55 mm long; cephalic sensilla setiform, 0.5–4 µm long; amphidial fovea sexually dimorphic, circular in female and doubleloop-shaped in male; oral opening located on the dorsal side of the body; secretory-excretory pore located at the level of cephalic setae bases; tail elongate-conoid; ovaries antidromously reflexed; testes opposed; spicules 16–21 µm long, curved, with weakly defined manubrium and subcylindrical shaft; gubernaculum plate-like, with caudal apophysis; supplements absent.
Material examined
SWEDEN: 2 ♂♂, 2 ♀♀, Skagerrak off the west coast of Sweden, 58°20′06″ N, 11°09′24″ E, muddy sediment from 53 m deep, 9 Aug. 2011, O. Holovachov leg. ( SMNH 161699–161701); 1 ♂, Skagerrak off the west coast of Sweden, 58°18′06″ N, 11°05′22″ E, mud and gravel from 40–60 m deep, 9 Aug. 2011, O. Holovachov leg. ( SMNH 161702); 1 ♂, Skagerrak off the west coast of Sweden, 58°22′14″ N, 11°05′00″ E, gravel, mud and algae from 30–70 m deep, 9 Aug. 2011, O. Holovachov leg. ( SMNH 161703); 3 ♂♂, 3 ♀♀, Skagerrak off the west coast of Sweden, 58°21′35″ N, 11°08′30″ E, fine gravel from 20–60 m deep, 9 Aug. 2011, O. Holovachov leg. ( SMNH 161704); 6 ♂♂, 9 ♀♀, Skagerrak off the west coast of Sweden, 58°7′58″ N, 11°10′05″ E, coarse gravel from 30–50 m deep, 9 Aug. 2011, O. Holovachov leg. ( SMNH 161705–161707).
Description
Adult
Body fusiform in female and cylindrical in male, tapering anteriorly in the pharyngeal region and posteriorly in the tail region, ventrally curved upon fixation. Cuticle finely striated along entire body, striae without longitudinal incisures or any other ornamentation visible under the light microscope. Somatic setae indistinct if present. Labial region rounded, lips fused. Inner and outer labial sensilla not observed. Cephalic sensilla small setiform, equal to 0.1–0.2 labial region diameters in length, their bases are located 3–7 µm from anterior end. Amphidial fovea dissimilar in shape and size between sexes (see descriptions of each sex). Oral opening shifted towards the dorsal side of the body, 1.5–3.5 µm from the anterior end. Stoma very small, cheilostom short, remaining part of stoma undifferentiated, its lining is uniform with the lining of the pharynx. Pharynx distinctly subdivided into anterior corpus and posterior postcorpus; corpus slightly fusiform, muscular; postcorpus narrow and long. Pharyngeal gland nuclei indistinct. Pharyngeal gland orifices penetrate pharyngeal lumen at the base of the stoma. Postcorpus is surrounded by six large pseudocoelomocytes, two cells arranged longitudinally in three body sectors: dorsal, right-subventral and left-subventral. Nerve ring is located at the posterior end of the pharynx, its exact position if obscured by small glandular cells (neuron bodies?). Secretory-excretory system present; renette cell located opposite to the ventral side of the anterior part of intestine; secretory-excretory pore opens to the exterior just posterior to cephalic setae bases. Tail elongate-conoid. Caudal glands and spinneret present.
Female
Amphidial fovea unispiral (circular). Reproductive system didelphic, amphidelphic; ovary branches symmetrical, antidromously reflexed. Anterior ovary situated to either right or left of intestine; posterior ovary situated to either right or left of intestine. Spermathecae present, filled with round spermathozoa. Vulva located at mid-body. Vagina straight, pars proximalis vaginae surrounded by a sphincter muscle; pars refringens vaginae absent. Intrauterine egg not seen. Rectum short.
Male
Amphidial fovea double-loop-shaped: starting from the porus amphidialis, fovea extends anteriorly for a short distance, makes a loop towards ventral side of the body and extends posteriorly for the full length of the amphid, makes another loop towards dorsal side of the body and extends anteriorly almost to the level of the porus amphidialis. Amphid length (longest) to width ratio is 2.7–5.2. Narrow space between amphidial branches not refractive and not ornamented. Reproductive system diorchic, anterior testis outstretched; posterior testis reflexed. Spicules paired and symmetrical, weakly curved, with weakly defined manubrium and subcylindrical shaft. Gubernaculum plate-like, with strong dorsocaudal apophysis. Supplements and caudal setae indistinct or absent.
Remarks
Both published descriptions of Diplopeltula belgica are each based on single male specimens ( Vincx & Gourbault 1992; Fadeeva & Mordukhovich 2013); females of this species were unknown until now. Recent male specimens resemble type male from the North Sea ( Vincx & Gourbault 1992) in structure of amphid, digestive (see above) and excretory systems, with the exception of the following: shorter body (358–418 µm in recent specimens vs 550 µm in type male), shorter amphid (13.5–18.5 µm in recent specimens vs 26 µm in type male), shorter cephalic setae (0.5–1.5 µm in recent specimens vs 4 µm in type male), shorter (16.5–18 µm in recent specimens vs 21 µm in type male) and less curved (weakly arcuate in recent specimens vs strongly arcuate in type male) spicules.
Specimen from the Sea of Japan ( Fadeeva & Mordukhovich 2013) is closer in size to specimens from Skagerrak ( Table 1 View Table 1 ) – the only substantial differences are the length of the amphidial fovea (13.5– 18.5 µm in recent specimens vs 37 µm in the male from the Sea of Japan) and less curved (weakly arcuate in recent specimens vs strongly arcuate in the male from the Sea of Japan) spicules.
Greatest discrepancies between all three populations are in the assumed position of the nerve ring and the morphology of male gonad. In the original description, the nerve ring is drawn short distance behind the amphid, at 54% of the pharynx length ( Vincx & Gourbault 1992). Male specimen from the Sea of Japan is described to have the nerve ring at 77% of the pharynx length ( Fadeeva & Mordukhovich 2013), but it is not drawn on the figure. In recent specimens, the position of the nerve ring can not be identified with confidence, and further observations are required to clarify this. Regarding the morphology of the male gonad, type specimen is said to have both testes outstretched ( Vincx & Gourbault 1992), while our observations clearly show that the posterior testis in this species is reflexed posteriad.
| SMNH |
Department of Paleozoology, Swedish Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |