
Akodon diauarum, Brandão & Carmignotto & Percequillo & Christoff & Mendes-Oliveira & Geise, 2022
|
publication ID |
https://doi.org/10.11646/zootaxa.5205.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:D3AADC46-80C5-4BCF-B0F5-6C074B42175F |
|
DOI |
https://doi.org/10.5281/zenodo.7320014 |
|
persistent identifier |
https://treatment.plazi.org/id/18673758-FF8B-2471-FEDF-FE697DA0984D |
|
treatment provided by |
Plazi |
|
scientific name |
Akodon diauarum |
| status |
sp. nov. |
Akodon diauarum , new species
( Figures 3–10 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
Diauarum Grass Mouse rato-do-mato-de-diauarum
Synonyms:
Akodon sp. : Silva & Yonenaga-Yassuda, 1998:47; Fagundes & Nogueira, 2007:1; Mendes-Oliveira, Carvalho Junior, Santos, Maria, Lima, Rossi, Saggin, Souza Junior & Dourado, 2010:144; Mendes-Oliveira, Santos, Carvalho Junior, Montag, Lima, Maria & Rossi, 2012:60.
Akodon sp. nov.: Lambert, Malcolm & Zimmerman, 2005a:984; Lambert, Malcolm & Zimmerman, 2005b:385; Lambert, Malcolm & Zimmerman, 2006:768.
Akodon sp. n.: Silva, Patton & Yonenaga-Yassuda, 2006:469; Ventura, O’Brien, Yonenaga-Yassuda & Ferguson-Smith, 2009:1; Di-Nizo, Banci, Sato-Kuwabara & Silva, 2017:838.
Akodon sp. (2n = 10): Geise, 2012:34; Pereira, Malcher, Nagamachi, O’Brien, Ferguson-Smith, Mendes-Oliveira & Pieczarka, 2016:2; Brandão, Garbino, Semedo, Feijó, Nascimento, Fernandes-Ferreira, Rossi, Dalponte & Carmignotto, 2019:281.
Akodon cf. cursor: Gettinger, Ardente & Martins-Hatano, 2012:154 View in CoL ; Ardente, Ferreguetti, Gettinger, Leal, Mendes-Oliveira, Martins-Hatano & Bergallo, 2016:6; Ardente, Ferreguetti, Gettinger, Leal, Martins-Hatano & Bergallo, 2017:128.
Akodon sp. 2 n = 10: Pardiñas, Teta, Alvarado-Serrano, Geise, Jayat, Ortiz, Gonçalves & D’Elía, 2015:203.
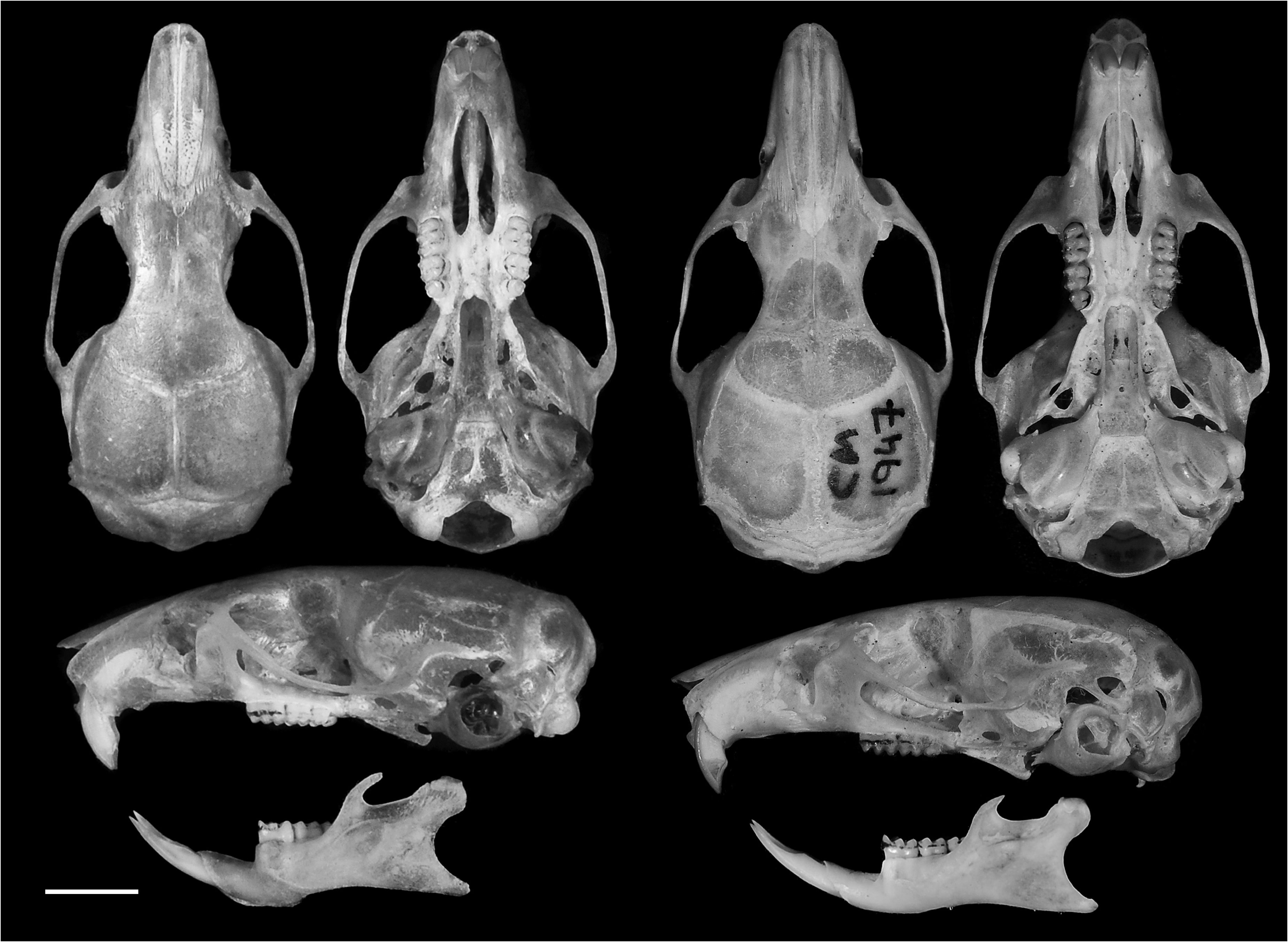
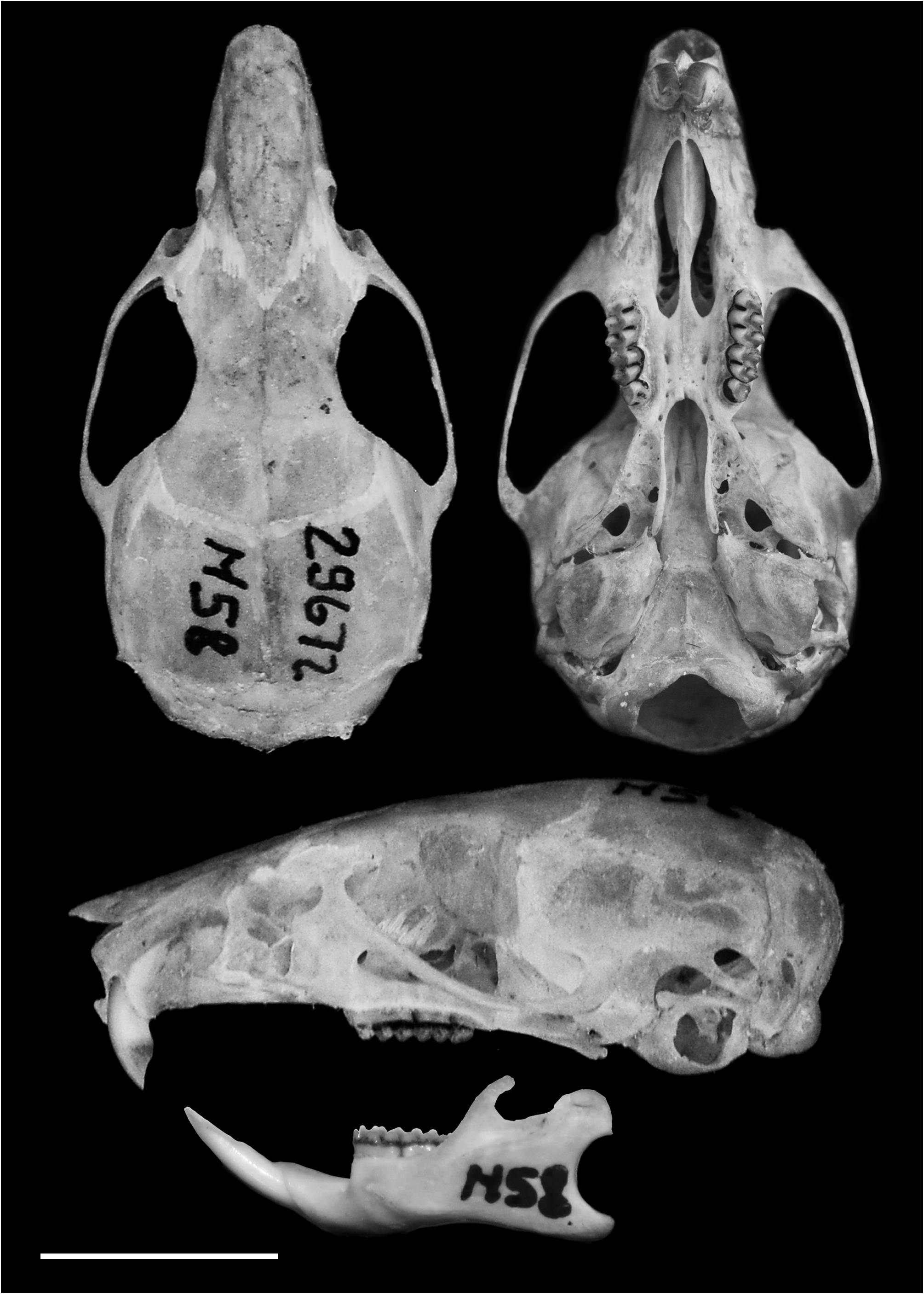
Holotype. MZUSP 29672 View Materials , adult male (age class 3), preserved as skin, skull ( Figure 7 View FIGURE 7 ), skeleton, and tissue in alcohol collected by A.P. Carmignotto, on May 8 th, 1997 (field number M58). Karyotype 2n=10, FN=16 (karyotype lamina number: CIT 575).
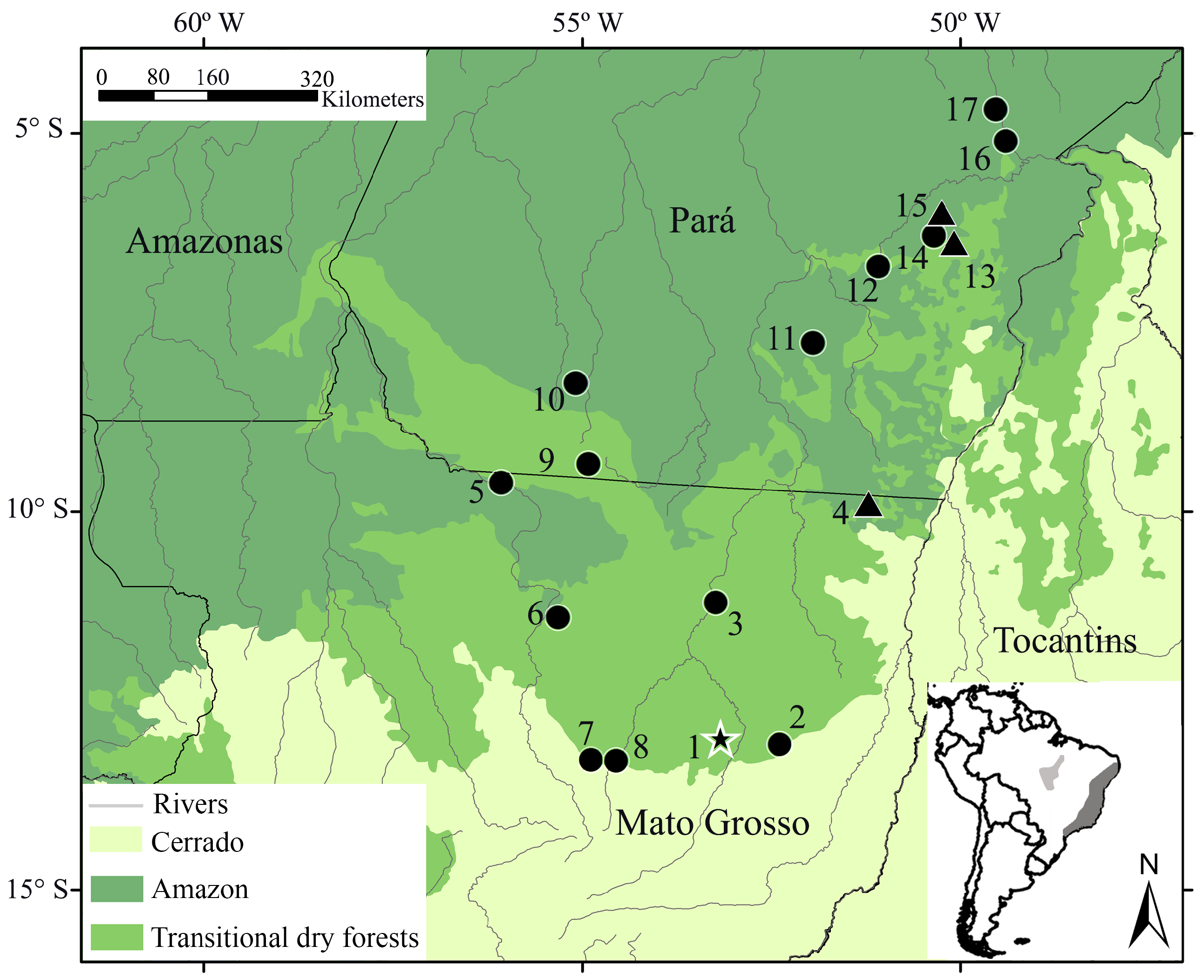
Type locality. 30 km to the north of the Gaúcha do Norte municipality, between the Coluene and Curisevo river basins, in northeastern Mato Grosso state, Brazil ( 13 o 00’53”S; 53 o 10’39”W [Datum SAD 69], 359 m. a.s.l.). This locality currently lies in the limits of the Mato Grosso seasonal forest ecoregion (see Dinerstein et al. 2017) located in transitional regions between the Amazonia and Cerrado domains ( Figure 1 View FIGURE 1 ) GoogleMaps .
Paratypes. MZUSP 29664 View Materials , 29674 View Materials , 29676 View Materials , 29677 View Materials , 29678 View Materials , 29683 View Materials ; MN 11691, 73906 , 73970 .
Diagnosis. This species is diagnosed by the following exclusive combination of traits: large-sized Akodon (HBL mean = 115.66 mm; CIL mean = 27.35 mm; LMR mean = 4.42 mm; see Table 9 View TABLE 9 ); tail shorter than head and body length (TL mean = 86.81 mm); overall pelage dark brown; eye-ring absent or inconspicuous; skull relatively robust, with moderately long and broad rostrum (LRO mean = 11.28 mm; ROW mean = 5.33 mm); moderately deep zygomatic notches; moderately broad zygomatic plate (BZP mean = 2.83 mm); interorbital region hourglass-shaped and with rounded supraorbital margins; long incisive foramina reaching the protocone of M1 (LFI mean = 6.56 mm); short palatal bridge ( sensu Hershkovitz 1962; LPB mean = 4.07 mm), with anterior margin of mesopterygoid fossa wide, with margins U-shaped (median palatine process absent) and even or slightly surpassing the posterior M3 level; sphenopalatine vacuities absent (or rarely restricted to small openings); parapterygoid fossae (at midlevel) approximately as wide as the mesopterygoid fossa; parietal conspicuously expanded onto lateral surface of braincase, with posterior margin in contact with exoccipital; frontosquamosal suture anterior to frontoparietal suture; mandible with moderate robust angular, condyloid and coronoid processes; upper incisors opisthodont; upper and lower molars moderately hypsodont with paraflexus and metaflexus well developed and deep in M1 and M2, nearly in a 90ºangle; and mesoflexid developed and deep in m1, m2 and m3, nearly in a 90ºangle; a unique chromosomal formula with 2n = 10 and FN =14-16.
Distribution. Known from the dry forest transition zone (Amazonia- Cerrado) of northeastern Mato Grosso, southern and southeastern Pará state, Brazil; in the southern limits of the Amazon biome ( Figure 1 View FIGURE 1 ). The altitudinal range of this species varies between 100 to 710 m. a.s.l.
Etymology. The specific epithet is a reference to Campo de Diauarum, a locality within the reserve Parque Indígena do Xingu (locality 3; Fig 1 View FIGURE 1 ). This reserve is the first indigenous land recognized in Brazil (in 1961) and is currently inhabited by 14 autochthonous ethnic groups. Campo de Diauarum is the locality where the oldest known voucher specimens (MN 11687, 11688, 11690-11694) of Akodon diauarum n. sp. were collected by the prestigious naturalist Helmut Sick during the Expedition Roncador-Xingu in 1949, under the leadership of the Villas Bôas brothers ( Sick 1997). This reserve is a highly preserved area that is still strongly resistant to the constant anthropic advance on southern Amazon, along the arc of deforestation ( Lima et al. 2011; Marques et al. 2020; Ferrante et al. 2021). Thus, we herein honor all the players of this scenario: the indigenous native people of the region, the Villas Bôas brothers, and Helmut Sick.
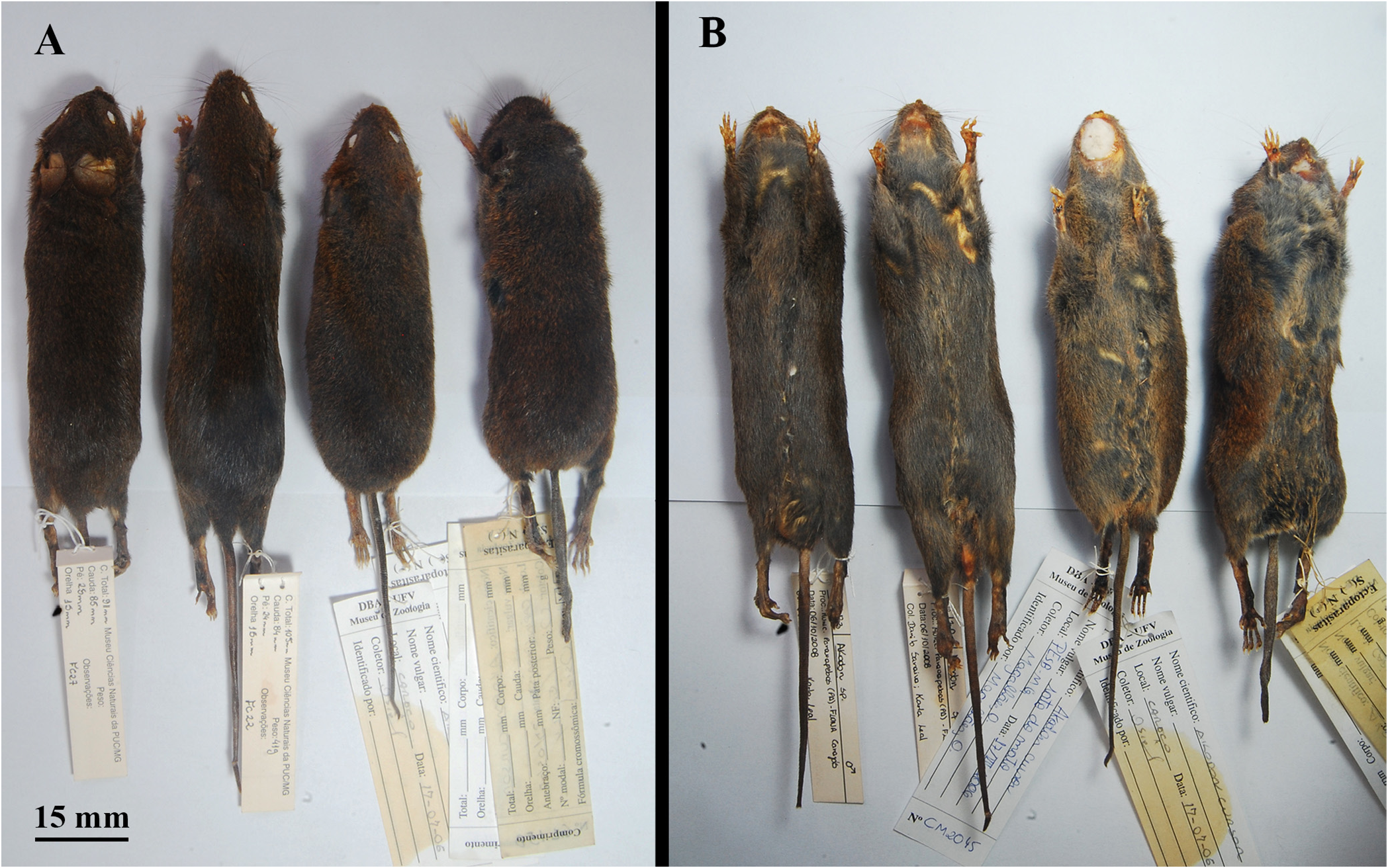
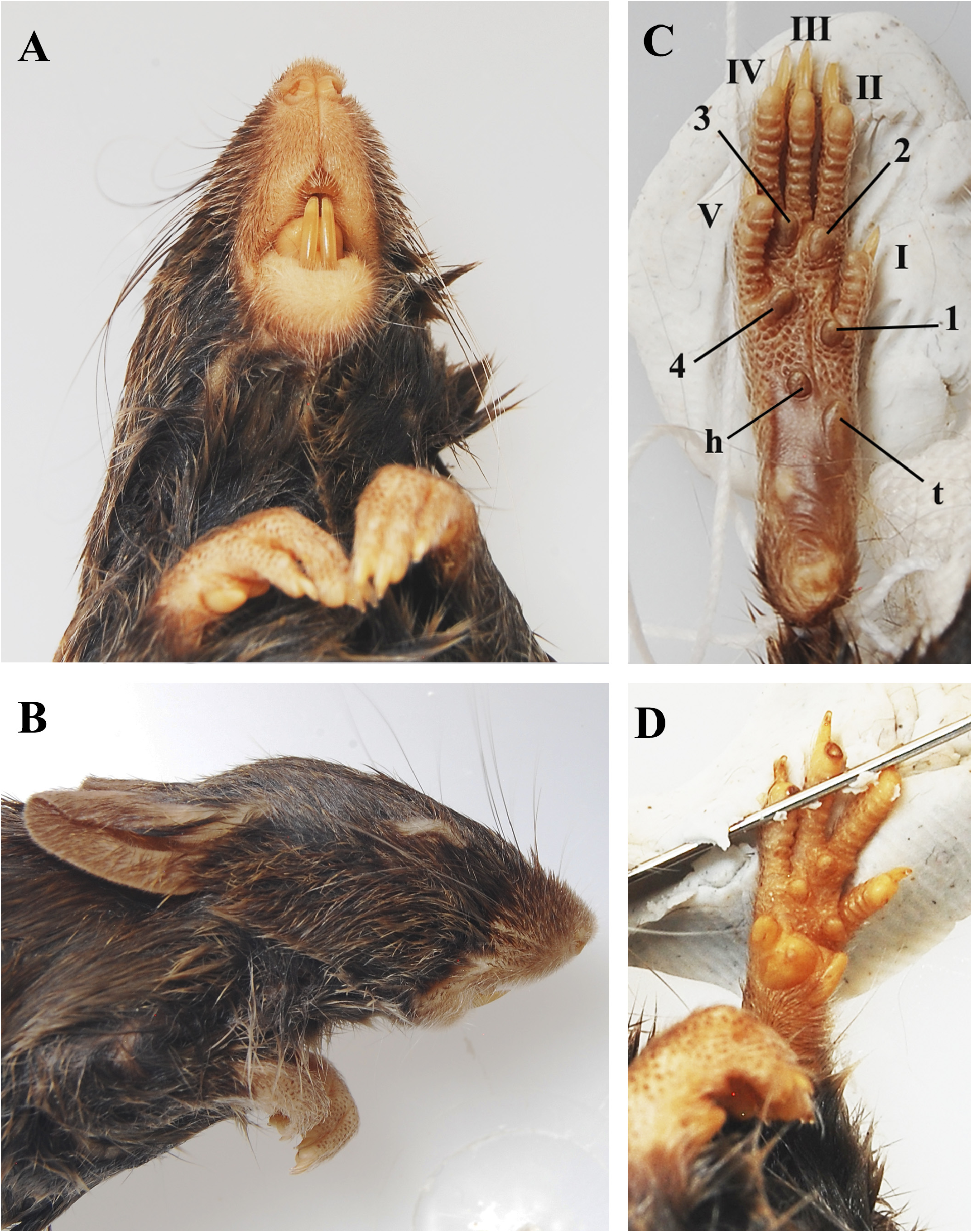
Morphological description. Large-sized Akodon , with a large head and body length (HBL mean = 115.66 mm) and body mass (mean = 46.99 g); tail length shorter than the HBL (mean of 73% of HBL); hind feet moderately narrow and long (mean of 23% of HBL); pinnae prominent (mean of 17% of HBL) ( Table 9 View TABLE 9 ). Overall pelage coloration dark brown, with gray-based hairs; dorsum covered by moderately long hairs (mean = 10 mm), slightly crisp, agouti and brown (from crown to rump); ventral region homogeneously dark grayish-brown, with a yellow/orangish wash mostly between forearms and at the ventral midline in some specimens; gradual transition between dorsal to ventral coloration, almost indistinguishable ( Figure 3 View FIGURE 3 ). Eye-ring inconspicuous ( Figure 8B View FIGURE 8 ), some specimens may exhibit a trace of it, but never as conspicuous as in other congeners (see Myers 1990). Mystacial vibrissae blackish and/ or brownish colored, and moderately long (reaching but not surpassing pinnae when laid back); ears covered by delicate and short brownish hairs ( Figure 8 View FIGURE 8 ). Tail unicolored (or inconspicuously darker above), covered with short and sparse blackish hairs dorsally and whitish hairs ventrally, scales small but visible to the naked eye ( Figure 3 View FIGURE 3 ), regular, and with 1-2 hairs per scale. Dorsal surface of manus and pes buffy-brown, sparsely covered by a mixture of silvery and brownish hairs, with claws moderate in length (mean = 2 mm), all covered by silvery and brownish ungual tufts slightly longer than claws; ventral surface naked and unpigmented; manus short and with two large carpals and three moderately large interdigital pads; pes relatively long and narrow, with outer digits (I and V) much shorter than the central ones (II, III and IV, all similar sized); ventral surface with distinct squamae (scalelike tubercles) well distributed, and with moderately large and fleshy interdigital (1-4) and tarsal pads (1 thenar and 1 hypothenar); all plantar pads are well developed, with only slightly different sizes: the thenar is the largest one; followed by interdigitals 2, 3 and 4, which are subequal in size; and interdigital 1 and hypothenar, being the smallest ones ( Figure 8 View FIGURE 8 ).
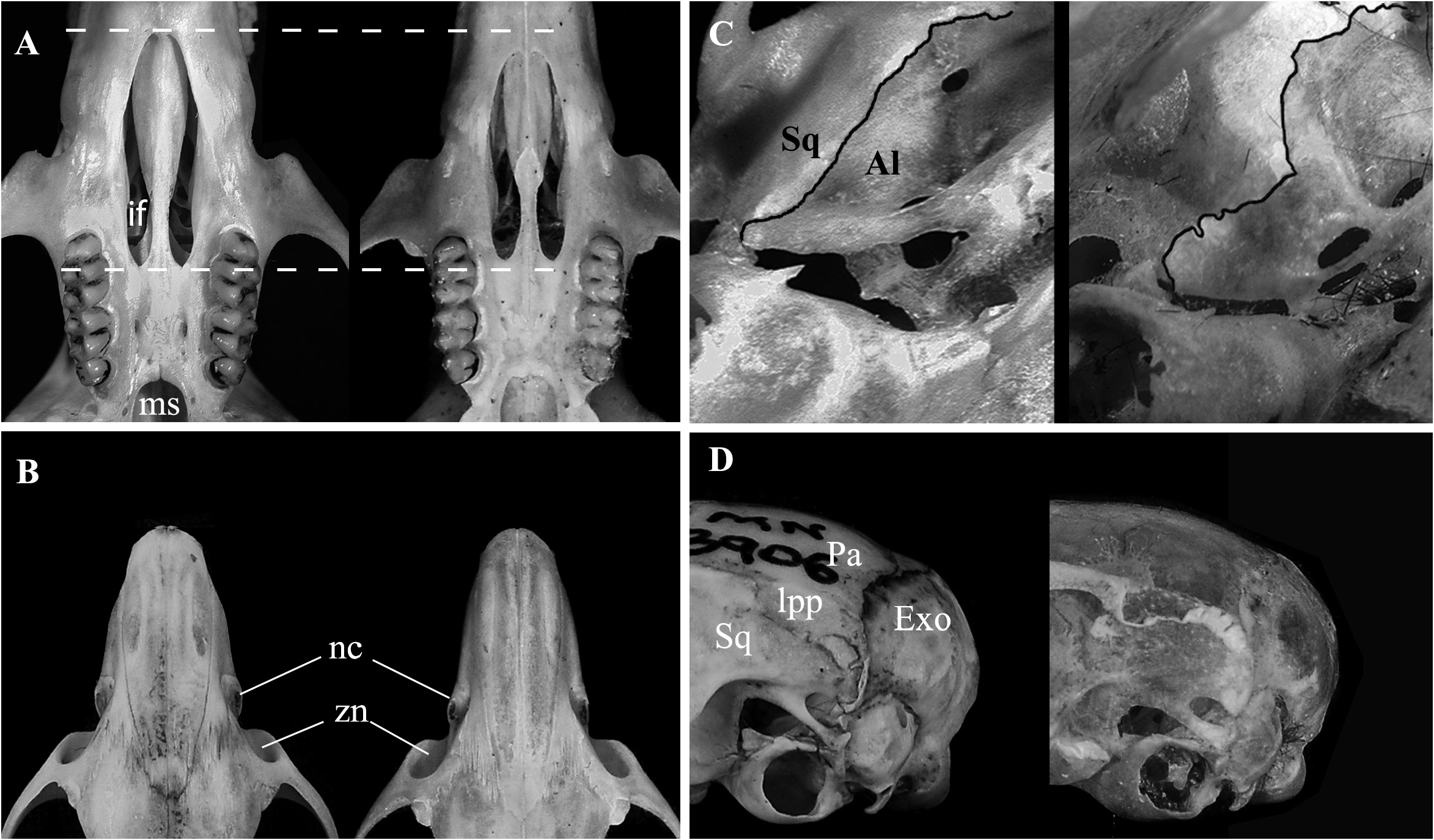
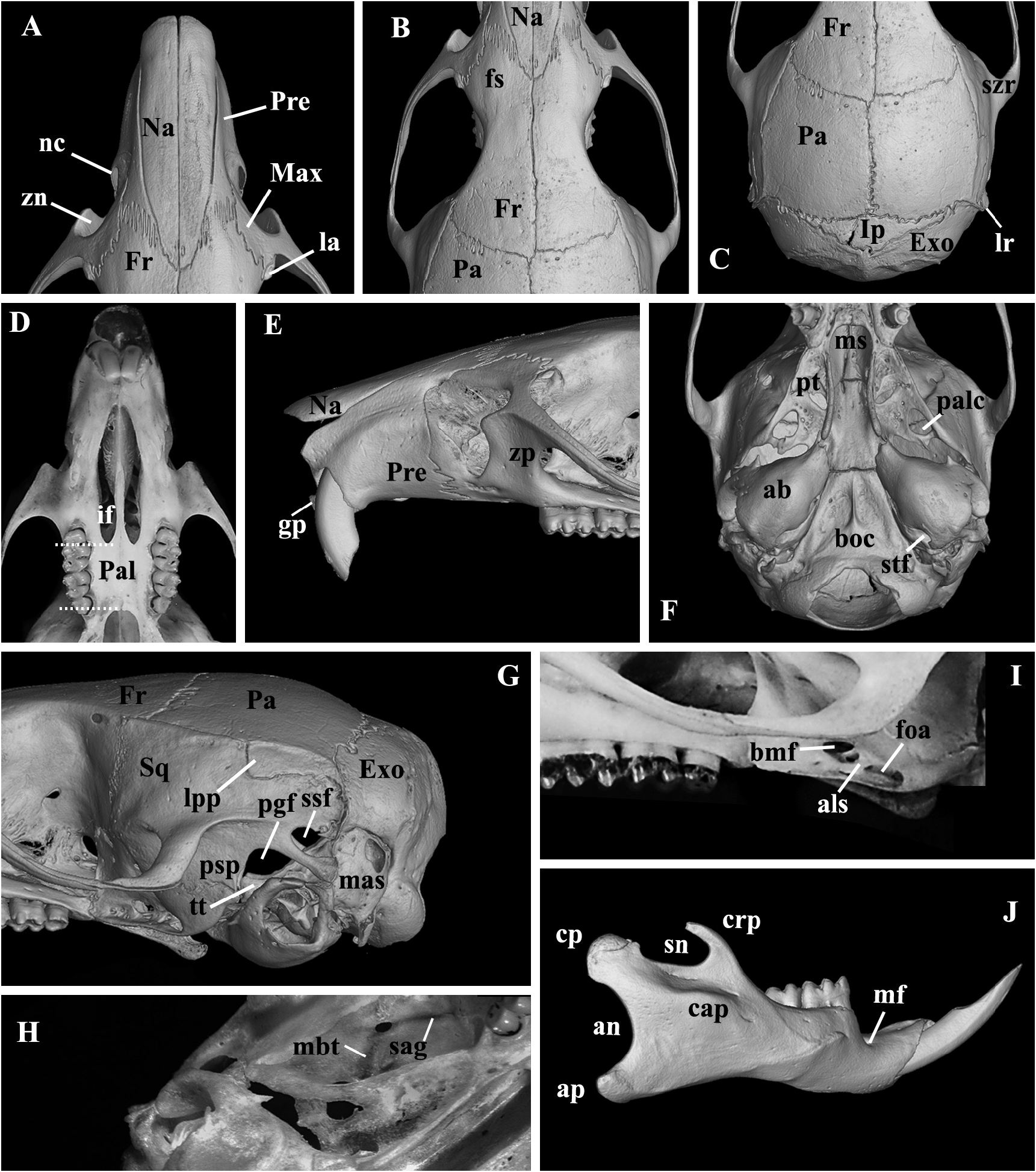
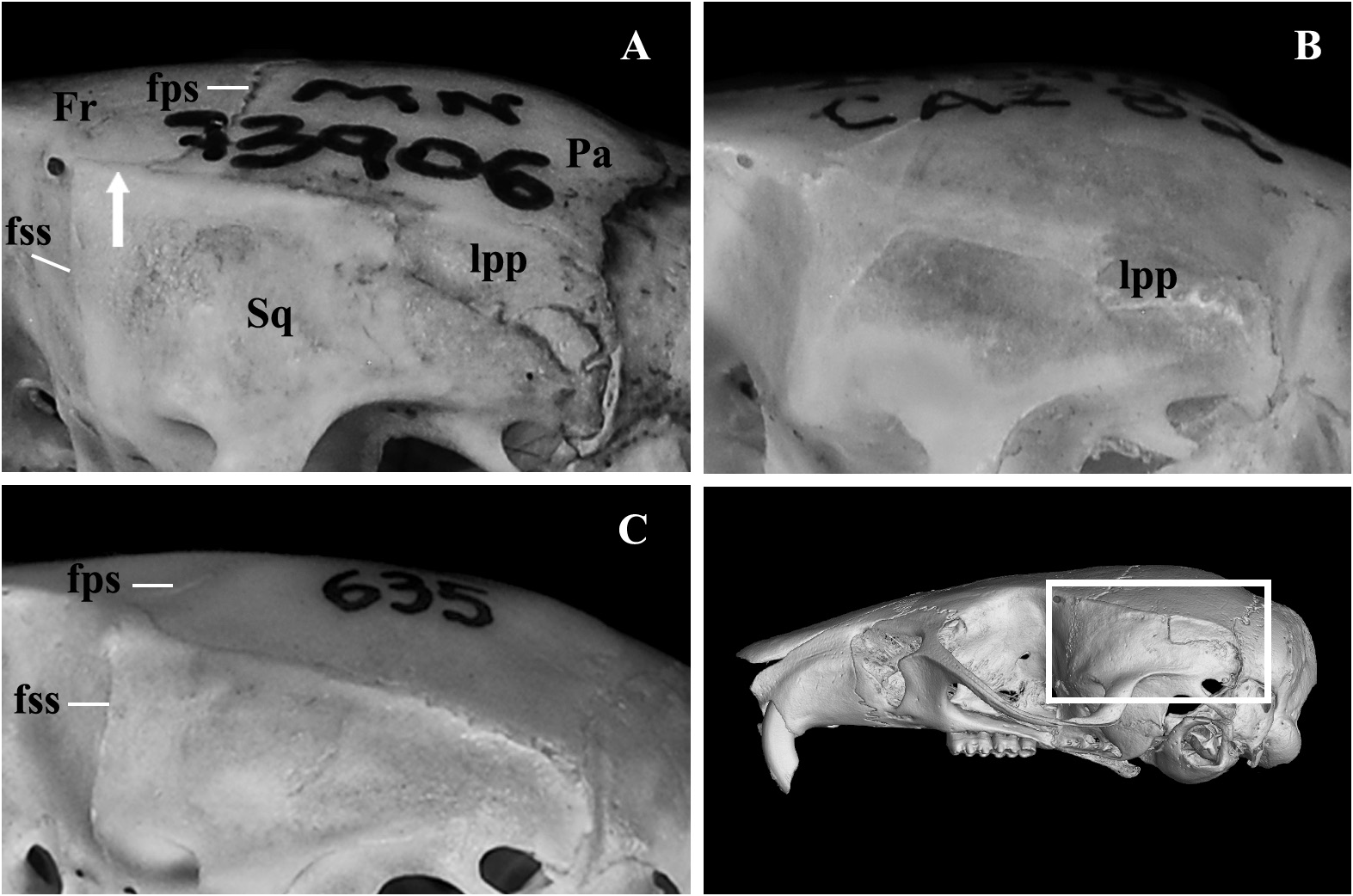
Skull (in dorsal view) characterized by a moderately long and broad rostrum (LRO mean = 11.28 mm; ROW mean = 5.33 mm) ( Figures 4 View FIGURE 4 , 9A View FIGURE 9 ); rostrum with a relatively developed capsular projection of nasolacrimal foramen and flanked by moderately deep zygomatic notches ( Figure 9A View FIGURE 9 ); nasals moderately wide, with sub-parallel margins that acuminate (U-shaped) only at the posterior end ( Figure 9A View FIGURE 9 ), and long, extending anteriorly beyond the premaxillae ( Figure 9E View FIGURE 9 ) and posteriorly near the lacrimal level ( Figure 9A View FIGURE 9 ); gnathic process short, slightly projecting anteriorly the upper incisors anterior surface ( Figure 9E View FIGURE 9 ). Nasofrontal ( Figure 9B View FIGURE 9 ) and frontoparietal ( Figure 9C View FIGURE 9 ) sutures V and U-shaped, respectively. Interorbital region hourglass-shaped, moderate in width (LIW mean = 5.12 mm), and with rounded supraorbital margins ( Figure 9B View FIGURE 9 ); frontal sinuses moderately inflated ( Figure 9B View FIGURE 9 ). Lacrimals moderately developed, with major and reduced contact with maxillary and frontal, respectively. Braincase oblong and squared (especially in dorsal view), with lambdoidal crests moderately developed ( Figure 9C View FIGURE 9 ) and temporal beads or ridges absent or poorly developed; interparietal narrow relative to its length, and with moderately long sutures between the parietal and exoccipital ( Figure 9C View FIGURE 9 ). Dorsal profile (in lateral view) slightly convex ( Figure 4 View FIGURE 4 ); premaxillae and nasals slightly projected anteriorly beyond the incisors ( Figure 9E View FIGURE 9 ); zygomatic plate relatively broad (BZP mean = 2.83 mm), with anterior margin straight in most individuals ( Figure 4 View FIGURE 4 ), only few individuals exhibit a concave margin ( Figure 9E View FIGURE 9 ), and with zygomatic spine absent; small masseteric scar present near the root of the zygomatic plate. Parietal conspicuously expanded onto lateral surface of braincase (posteriorly to the end of the squamosal zygomatic root), frequently in contact to exoccipital or the lambdoidal crests ( Figure 9G View FIGURE 9 ); frontosquamosal suture anterior to frontoparietal suture ( Figure 9G View FIGURE 9 ), which allows further contact between the squamosal and frontal. Zygomatic arch moderately developed (formed by jugal, zygomatic process of maxillary and zygomatic process of squamosal); zygomatic arches breadth (ZIB mean = 14.67 mm) conspicuously wider than braincase (BBR mean = 11.51 mm) ( Figures 4 View FIGURE 4 , 9C View FIGURE 9 ). Stapedial foramen, sphenofrontal foramen, and squamosal-alisphenoid groove present, configuring the primitive carotid circulatory pattern 1 ( sensu Voss 1988) ( Figures 9F, 9H View FIGURE 9 ). Alisphenoid strut present in almost all specimens configuring discrete oval accessory and buccinator-masticatory foramina ( Figure 9I View FIGURE 9 ). Hamular process of squamosal wide, resulting in a moderately developed postglenoid foramen and subsquamosal fenestra, being the former larger; tegmen tympani overlapped to a small posterior suspensory process of squamosal; mastoid with a conspicuous fenestra ( Figure 9G View FIGURE 9 ). Incisive foramina moderately wide (BIF mean = 2.32 mm, wider near the premaxillae/maxillary suture) and long (LIF mean = 6.56 mm, averaging about 35.4% of diastema length, LDI mean = 7.90 mm), with posterior margins surpassing M1 protocone ( Figure 9D View FIGURE 9 ), with few specimens nearly reaching the hypoflexus level. Palate short and wide ( sensu Hershkovitz 1962), with anterior margin of mesopterygoid fossa aligned to or slightly surpassing the posterior margin of M3 ( Figure 9D View FIGURE 9 ); palate smooth, without pronounced traits, exhibiting shallow posterolateral pits mostly in older individuals. Mesopterygoid fossa wide ( Figure 9F View FIGURE 9 ), with anterior margin rounded and never with a median palatine process; bony roof of mesopterygoid fossa totally ossified or with inconspicuous sphenopalatine vacuities ( Figure 9F View FIGURE 9 ). Hamular process of pterygoid medium in size and slightly divergent at posterior end; parapterygoid fossa (at mid-level) approximately as wide as the mesopterygoid fossa; and with margins conspicuously straight and divergent posteriorly ( Figure 9F View FIGURE 9 ). Basioccipital wide and moderately excavated; auditory bullae flask-shaped and medium-sized, with long and wide eustachian tube ( Figure 9F View FIGURE 9 ). Foramen magnum oval in shape; occipital condyles medium-sized.
Mandible similar to congeners, but with somewhat robust angular, condyloid and coronoid processes ( Figure 9J View FIGURE 9 ). Coronoid process falciform, subequal in height to the condyloid process; angular process short, reaching the same level of condyloid process posteriorly. Sigmoid and angular notches moderately deep. Masseteric crest with superior and inferior ridges converging anteriorly as open chevron, which ends below the middle, or the procingulum, of ml level. Capsular process of lower incisors poorly developed, lying at the base of coronoid process; mental foramen visible in lateral view; anterior point of diastema located slightly below the alveolar plane.
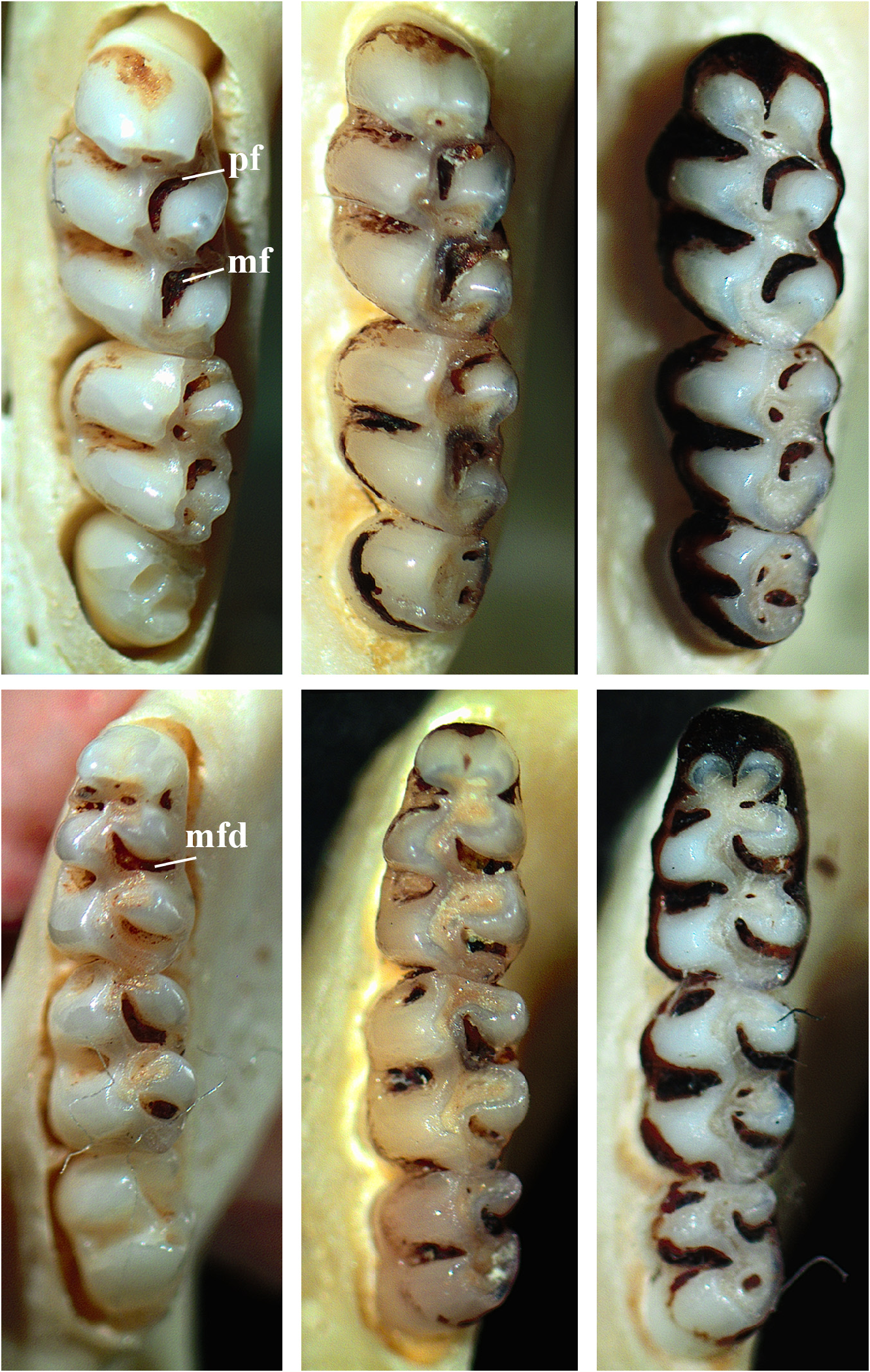
Upper and lower incisors opisthodont, without grooves, and with enamel light orange. Upper molar rows parallel; main cusps arranged in slightly alternated pairs, with protocone and hypocone slightly anterior to opposite cusps (paracone and metacone, respectively); labial (paracone and metacone) cusps slightly larger than the lingual (protocone and hypocone) cusps; molars crested ( sensu Hershkovitz 1962), narrow and elongated; upper and lower molars moderately hypsodont, M1 with inconspicuous anteroflexus and mesoflexus, either obliterated due to minimal wear or remaining as a small internal fossette, and conspicuous protoflexus, hypoflexus, paraflexus, and metaflexus, with the last two almost forming a 90º angle; procingulum of M1 separated by anteromedian flexus, resulting in an anterolabial and anterolingual conules subequal in size; anteroloph and parastyle present, but anteroloph coalesced to the anterolabial conule with early wear (thus, the anteroflexus is almost indistinguishable); protostyle absent; mesoloph reduced in length and generally coalesced with paracone due to wear; enteroloph and enterostyle absent; posteroloph present and with evidence of posteroflexus in young specimens, but both are poorly defined in subadults/adults due to early wear. M2 similar to M1; rectangular in outline, with anteroloph moderately developed; mesoloph very reduced and partially fused to paracone; flexus are just as developed, including a trace of a protoflexus and posteroflexus in young specimens. M3 half the size of M2, cylindrical in outline, with main cusps and flexus distinct in young specimens or mostly fused in subadult/adults due to early wear, but with paraflexus and metaflexus often remaining as a small internal fossette in older specimens ( Figure 6 View FIGURE 6 ).
Lower molars crested; labial cusps (protoconid and hypoconid) arranged in alternated pairs in relation to lingual (metaconid and entoconid) cusps, with metaconid and entoconid distinctly anterior to protoconid and hypoconid, respectively; m1 with anterolabial and anterolingual conulids of equal size, separated by a consistently present and deep anteromedian flexid; m1 with inconspicuous metaflexid and entoflexid, either obliterated due to minimal wear or remaining as a small internal fossette, and conspicuous protoflexid, hypoflexid, posteroflexid, and mesoflexid, with the last two almost forming a 90º angle; small protolophid present, connected to anterior murid by enamel lophid and fused with protostylid; anterolophid and metastylid absent; mesolophid very reduced (partially coalesced to entoconid due to wear) and fused to mesostylid; ectolophid absent; ectostylid absent or very reduced; posterolophid conspicuous and separated from entoconid by well-defined posteroflexid; m2 similar to m1, protolophid reduced, often fused with protoconid due to wear; flexids are just as developed, except for protoflexid; m3 greater in size than M3, with the four main cusps visible, mesolophid absent or very tiny; posterolophid might be present, but is probably fused to other cusps; protoflexid and posteroflexid mostly fused to cusps in mature adults; hypoflexid and mesoflexid fused only in very old specimens; entoflexid inconspicuous ( Figure 6 View FIGURE 6 ).
Morphological variation. Few specimens present discrete morphological variation: three specimens from Nova Ubiratã (locality 7, Figure 1 View FIGURE 1 ) present a white apical part of the tail, restricted to only about 10-20 mm ( UFMT BRM 165 , 175 , 279 ) ; in a few specimens the alisphenoid strut is absent ( MN 73906 , UFMT 3423), restricted to only one side of the skull ( UFMT 3476 , 4110 ) , or is notably thin in one ( UFMT BRM 165 ) or both sides ( UFMT BRM 175 ) ; and the anteromedian flexid is absent in one specimen ( UFMT BRM 164 ) .
Comparisons. Besides A. cursor , we also compared A. diauarum n. sp. to other members of the Akodon cursor species group. Akodon lindberghi and A. mystax are the most distinctive ones, since both are the smallest taxa, with ranges of CIL (20.69–22.88 mm, 21.07–23.18 mm, respectively) and LMR (3.52–3.94 mm, 3.54–3.93 mm) distinctly smaller than A. diauarum n. sp. (CIL 24.61–29.76 mm; LMR 3.94–5.35 mm— Tables 9 View TABLE 9 and S 1 View TABLE 1 ). In turn, A. montensis , A. paranaensis and A. reigi are similar-sized to A. diauarum n. sp., overlapping in most measurements ( Tables 9 View TABLE 9 and S 1 View TABLE 1 ; Geise et al. 2005; Gonçalves et al. 2007).
Qualitative characters of A. diauarum n. sp. that allow distinction to the aforementioned similar-sized species are: i) the conspicuously expanded lateral projection of parietal (lpp), usually in contact to the exoccipital versus lpp less developed, usually not in contact to the exoccipital ( A. paranaensis ) or absent (or inconspicuous, when present), exhibiting a straight lateral border of parietal ( A. montensis ); ii) frontosquamosal suture notably anterior to frontoparietal suture, which allows contact between the frontal and the dorsal surface of squamosal versus frontosquamosal suture only slightly anterior to the frontoparietal suture ( A. montensis , A. paranaensis ), with frontal and dorsal surface of squamosal not or only slightly in contact ( Figure 10 View FIGURE 10 ). Unfortunately, we did not have access to specimens of A. reigi . Nonetheless, this species is remarkably similar in morphometric dimensions and qualitative characters to Akodon paranaensis ( Gonçalves et al. 2007; Pardiñas et al. 2015), for which we made a close comparison to a large number of specimens (n = 37) of the type series (see Christoff et al. 2000). Moreover, the phylogenetic evidence based on cytb data, recovered these species as part of a monophyletic assemblage presenting similar karyotypes, of 2n = 44, FN = 44 ( Gonçalves et al. 2007; Coyner et al. 2013), being quite distinct to Akodon diauarum n. sp. (2n = 10, FN = 14-16) and its sister species A. cursor (2n = 14-16, FN = 18-26)—see further discussion below.
Karyological data. Chromosomal formula typically 2n = 10; FN = 14-16 ( Table 1 View TABLE 1 ), with one female with 2n = 9 due to monosomy of the X chromosome ( Silva & Yonenaga-Yassuda 1998).
Natural history. Specimens were captured during the wet and dry seasons, most by live traps (always at ground level), but also with pitfall traps ( Lambert et al. 2006; Ardente et al. 2017; A.P. Carmignotto field data). Males with scrotal testes were recorded in February, March, May, August and September; while pregnant females were recorded in January, February and May, either with two or four embryos. One adult female has ten mammae in five pairs, one inguinal, one abdominal, one postaxial, one pectoral and one thoracic. Other simodontinae captured in the same geographic region of A. diauarum n. sp. include: Calomys tener , Calomys tocantinsi , Euryoryzomys emmonsae , Hylaeamys megacephalus , Neacomys amoenus , Neacomys xingu , Necromys lasiurus , Nectomys rattus , Neusticomys ferreirai , Oecomys bicolor , Oecomys catherinae , Oecomys roberti , Oligoryzomys mattogrossae , Oligoryzomys microtis , Oxymycterus amazonicus , Pseudoryzomys simplex , Rhipidomys emiliae ( Lambert et al. 2005a, b; Ardente et al. 2016; 2017; Suárez-Villota et al. 2018; Brandão et al. 2019; Guilard et al. 2020; A.P. Carmingotto pers. communication).
Akodon diauarum n. sp. occurs mainly in seasonal deciduous forests in the transition zone between the Amazon and the Cerrado, in the southern limits of the Amazon biome. At Serra dos Carajás (locality 15, Figure 1 View FIGURE 1 ) it was also recorded in areas of metallophilic savanna or “canga”, occurring in much higher abundance than in adjacent forests ( Ardente et al. 2016). The “canga” is characterized by natural enclaves of rocky vegetation in the middle of the forest, with herbaceous and arboreal components that grow above the rocky substrate with iron ore deposits, being a unique ecosystem of plants, adapted to local environmental conditions, including several endemic species ( Viana & Mota 2016).
This grass mouse seems to be abundant in some localities, ranking as the most abundant species in Gaúcha do Norte (A.P. Carmignotto, field data; locality 1– Figure 1 View FIGURE 1 ), and one of the most abundant in other localities ( Lambert et al. 2005a; Ardente et al. 2016; localities 11 and 15– Figure 1 View FIGURE 1 ). Nonetheless, was relatively rare in others, such as Vila Rica (A.P. Carmignotto, field data; locality 4– Figure 1 View FIGURE 1 ) and Tanguro Farm ( Mendes-Oliveira et al. 2012; locality 2– Figure 1 View FIGURE 1 ). Current data indicate that the species present some degree of tolerance to human disturbance. Ardente et al. (2017) reported high detectability of A. diauarum n. sp. in impacted mining areas, Lambert et al. (2005b) indicated that the abundance of this species was higher in edge-affected or disturbed habitats, and MendesOliveira et al. (2012) captured the species only at forest edges. Together with its preference to canga, these data suggest that this species is associated with more open and transitional formations, including disturbed areas, an ecological feature similar to other species of the genus Akodon , including other taxa of the A. cursor species group ( Umetsu & Pardini 2007; Queirolo & Granzinolli 2009; Geise 2012; Pardiñas et al. 2015).
Although stomach contents or feeding behavior are not known for the species, Lambert et al. (2005b) suggest that it likely feeds on insects and seeds, reporting that A. diauarum n. sp. is associated with areas of seed predation of mahogany ( Swietenia macrophylla ) and areas with high insect biomass. Ardente et al. (2017) captured A. diauarum n. sp. with the use of three different types of bait: bacon, banana and peanut butter, resulting in distinct capture success (16%, 26% and 42%, respectively), suggesting its preference for plant-based itens.
| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
| UFMT |
Universidade Federal de Mato Grosso |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Sigmodontinae |
|
Genus |
Akodon diauarum
| Brandão, Marcus Vinicius, Carmignotto, Ana Paula, Percequillo, Alexandre Reis, Christoff, Alexandre Uarth, Mendes-Oliveira, Ana Cristina & Geise, Lena 2022 |
Akodon sp. 2
| Pardinas, U. F. J. & Teta, P. & Alvarado-Serrano, D. F. & Geise, L. & Jayat, P. & Ortiz, P. E. & Goncalves, P. R. G. & D'Elia, G. 2015: 203 |
Akodon sp.
| Brandao, M. V. & Garbino, G. S. T. & Semedo, T. B. F. & Feijo, A. & Nascimento, F. O. & Fernandes-Ferreira, H. & Rossi, R. V. & Dalponte, J. & Carmignotto, A. P. 2019: 281 |
| Pereira, A. L. & Malcher, S. M. & Nagamachi, C. Y. & O'Brien, P. C. M. & Ferguson-Smith, M. A. & Mendes-Oliveira, A. C. & Pieczarka, J. C. 2016: 2 |
| Geise, L. 2012: 34 |
Akodon cf. cursor : Gettinger, Ardente & Martins-Hatano, 2012:154
| Gettinger, D. & Ardente, N. C. & Martins-Hatano, F. 2012: 154 |
Akodon sp.
| Di-Nizo, C. B. & Banci, K. R. S. & Sato-Kuwabara, Y. & Silva, M. J. J. 2017: 838 |
| Silva, M. J. J. & Patton, J. L. & Yonenaga-Yassuda, Y. 2006: 469 |
Akodon sp.
| Lambert, T. D. & Malcolm, J. R. & Zimmerman, B. L. 2006: 768 |
| Lambert, T. D. & Malcolm, J. R. & Zimmerman, B. L. 2005: 984 |
| Lambert, T. D. & Malcolm, J. R. & Zimmerman, B. L. 2005: 385 |
Akodon sp.
| Mendes-Oliveira, A. C. & Carvalho Junior, O. & Santos, P. G. P. & Maria, S. L. & Lima, R. C. S. & Rossi, R. V. & Saggin, P. G. & Souza Junior, A. P. & Dourado, G. 2010: 144 |
| Fagundes, V. & Nogueira, C. D. A. 2007: 1 |
| Silva, M. J. & Yonenaga-Yassuda, Y. 1998: 47 |