
Macrobiotus polypiformis, Roszkowska & Ostrowska & Stec & Janko & Kaczmarek, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.327 |
|
publication LSID |
lsid:zoobank.org:pub:C8F064E6-137C-4D50-A738-CCC828281CD1 |
|
DOI |
https://doi.org/10.5281/zenodo.3847980 |
|
persistent identifier |
https://treatment.plazi.org/id/3858645A-2909-4DB9-82CD-CF2B4D7051C2 |
|
taxon LSID |
lsid:zoobank.org:act:3858645A-2909-4DB9-82CD-CF2B4D7051C2 |
|
treatment provided by |
Carolina |
|
scientific name |
Macrobiotus polypiformis |
| status |
sp. nov. |
Macrobiotus polypiformis View in CoL sp. nov.
urn:lsid:zoobank.org:act:3858645A-2909-4DB9-82CD-CF2B4D7051C2
Figs 1–29 View Figs 1–2 View Figs 3–6 View Figs 7–9 View Figs 10–15 View Figs 16–19 View Figs 20–23 View Figs 24–29 ; Tables 3–4 View Table 3 View Table 4
Etymology
The specific epithet ‘ polypiformis ’ refers to the similarity of the egg processes to the polyp form found in the phylum Cnidaria.
Material examined
Specimens mounted on microscope slides in Hoyer’s medium, fixed on SEM stubs or processed for DNA sequencing.
Holotype
ECUADOR: slide 1215/ 19, 3 Jan. 2015, Milena Roszkowska and Łukasz Kaczmarek leg. ( MECN).
Paratypes
ECUADOR: 135 animals (including 10 simplex), one exuvia and 44 eggs (including two with developed embryos), same data as holotype ( MECN, slide 1215 /19 (with holotype), 4 paratypes (slides: 1215 /*, where the asterisk can be substituted by any of the following numbers: 23, 25) and two eggs (slides: 1215 /*: 8, 9); DATE, 92 paratypes (slides: 1215 /*: 1, 2, 3, 10, 11, 12, 13, 14, 15, 16, 19, 20, 22, 24) and 24 eggs (slides: 1215 /*: 1, 4, 5, 7, 26, 27); ZMUC, 5 paratypes (slides 1215 /*: 17, 21) and 3 eggs (slide 1215 /6)).
Type locality
ECUADOR: Manabí Province, 1°04′06″ S, 89°52′18″ W; 370 m asl, moss sample from a concrete wall, next to E15 road, ca 3.5 km W from San Lorenzo, tropical rainforest.
Description
Animals (measurements and statistics in Table 3 View Table 3 )
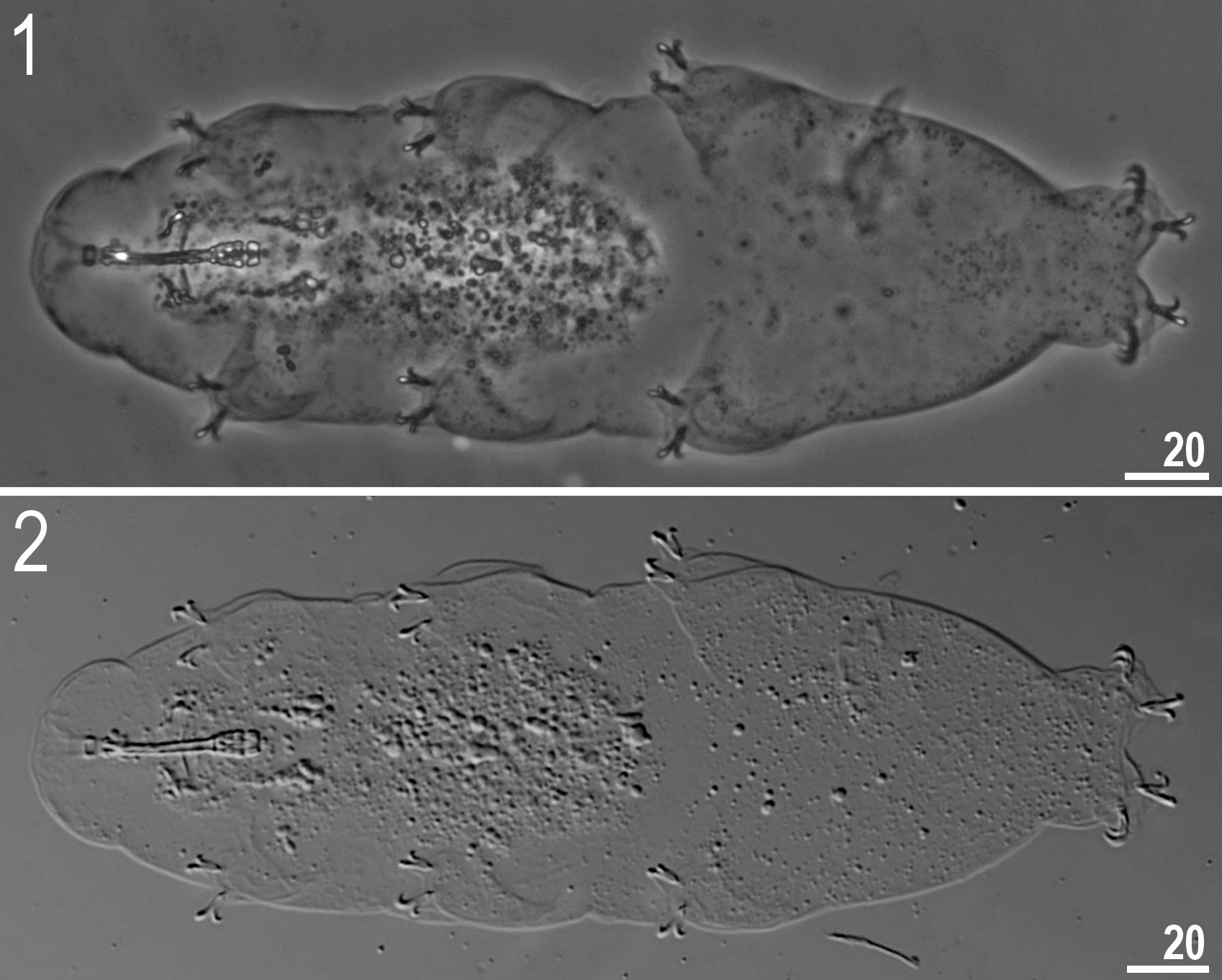
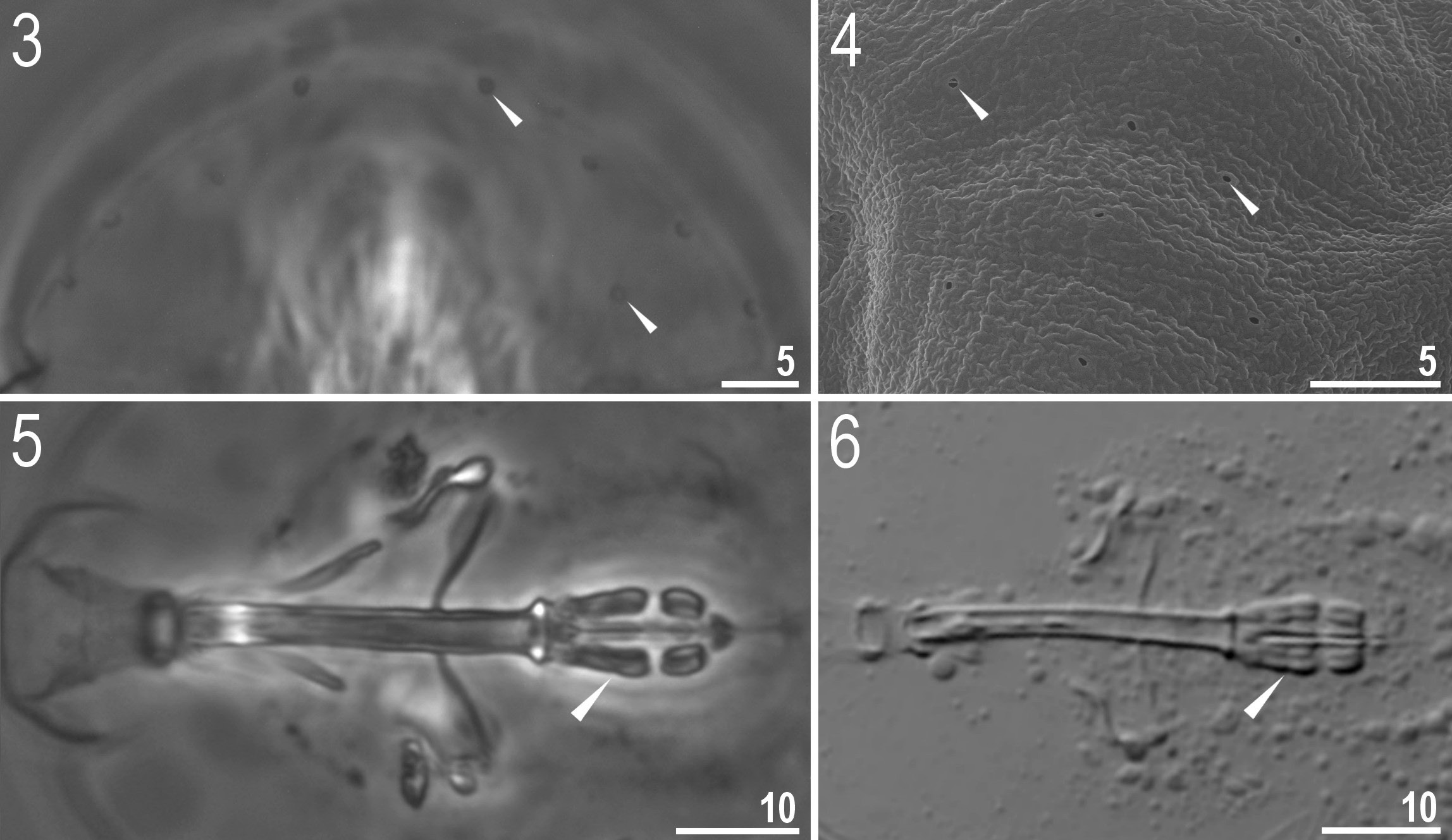
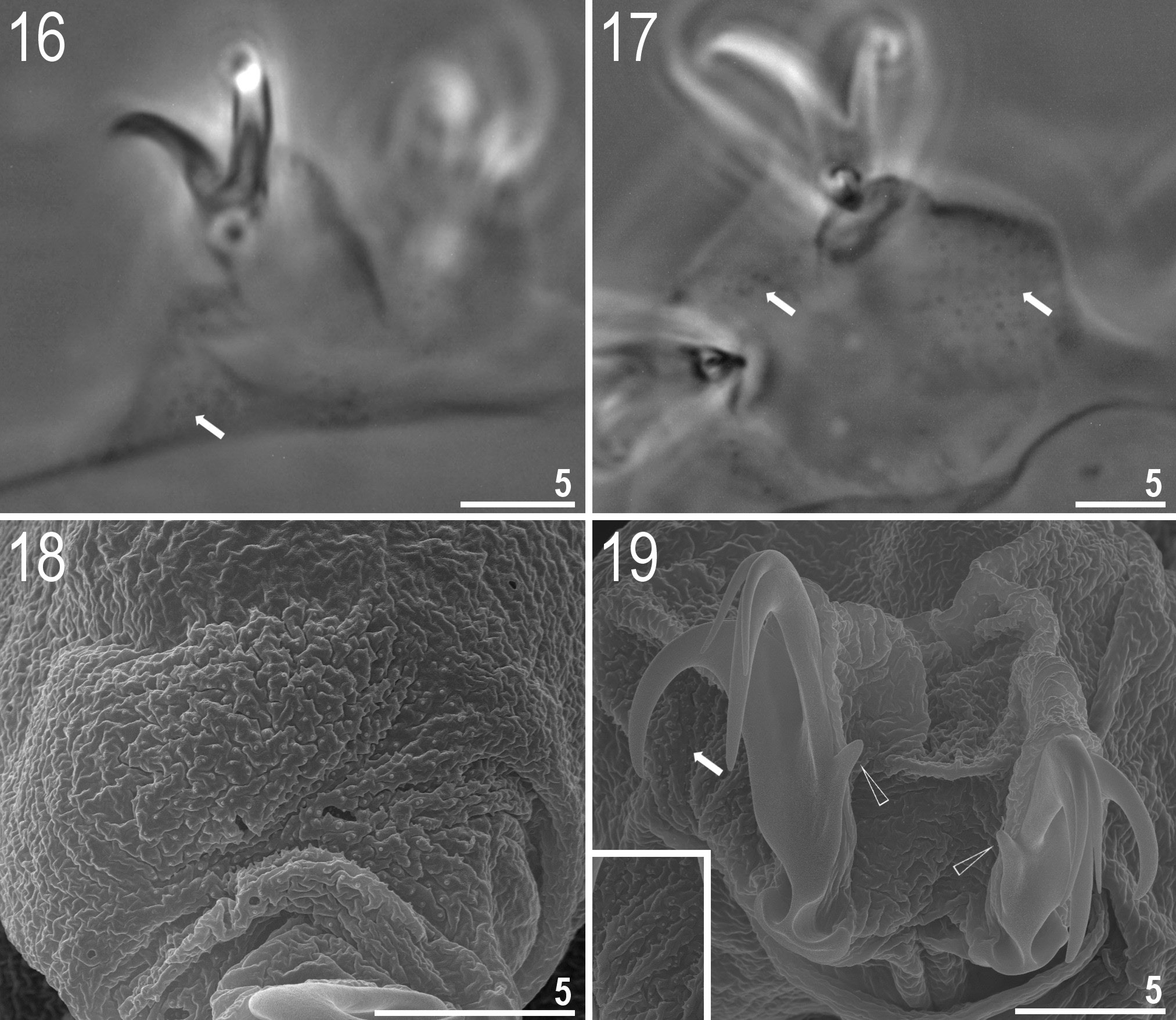
Body white in juveniles and adults, transparent after fixation in Hoyer’s medium ( Figs 1–2 View Figs 1–2 ). Eyes present (in 93% of measured specimens). Dorsal and ventral cuticle smooth under LM. Additionally, small oval and round pores (0.9–1.2 μm in diameter), sometimes difficult to observe under LM, are scattered randomly on the entire cuticle ( Figs 3–4 View Figs 3–6 ). A ring of pores, difficult to observe under LM, is present around the mouth opening, below the peribuccal sensory lobes. One patch of fine and dense granulation above claws on legs I–IV present ( Figs 16–19 View Figs 16–19 ).
Mouth antero-ventral. Bucco-pharyngeal apparatus of the Macrobiotus type ( Figs 5–6 View Figs 3–6 ), with the ventral lamina and ten small peribuccal lamellae ( Figs 6 View Figs 3–6 , 9 View Figs 7–9 ) followed by six buccal sensory lobes. Oral cavity armature composed of three bands of teeth, of which only the third band is visible under LM ( Fig. 7 View Figs 7–9 , empty arrowhead). SEM is required to reveal the first and the second bands of teeth (see Figs 8–9 View Figs 7–9 ). The first band of teeth comprises extremely small cones arranged in 3–4 rows situated at the anterior portion of the oral cavity, at the base of the peribuccal lamellae ( Figs 8–9 View Figs 7–9 , filled arrowheads). The second band of teeth is composed of 4–5 rows of small cones (but larger than those on the first band), positioned towards the rear of the oral cavity, between the ring fold and the third band of teeth ( Figs 8–9 View Figs 7–9 , arrows). The teeth of the third band are positioned at the rear of the oral cavity, between the second band of teeth and the buccal tube opening ( Figs 7–9 View Figs 7–9 , empty arrowheads).
Under LM, the teeth of the third band appear as a single, thin, transversal ridge both ventrally and dorsally ( Fig. 7 View Figs 7–9 , empty arrowhead). Although SEM reveals that both ventral and dorsal teeth do indeed form continuous ridges, there are evident median (M) and lateral (L) peaks corresponding to the median and lateral teeth found in species with better developed oral cavity armatures ( Figs 8–9 View Figs 7–9 ). Median teeth are smaller than the lateral teeth ( Fig. 8 View Figs 7–9 ). In addition, there are a number of smaller accessory teeth (a) placed laterally to the lateral teeth. These accessory teeth are better developed ventrally than dorsally ( Fig. 8 View Figs 7–9 ). Pharyngeal bulb spherical, with triangular apophyses, two rod-shaped macroplacoids and a triangular microplacoid ( Figs 5–6 View Figs 3–6 ). The macroplacoid length sequence 1>2. The first macroplacoid with a central constriction ( Figs 5–6 View Figs 3–6 , the filled arrowhead).
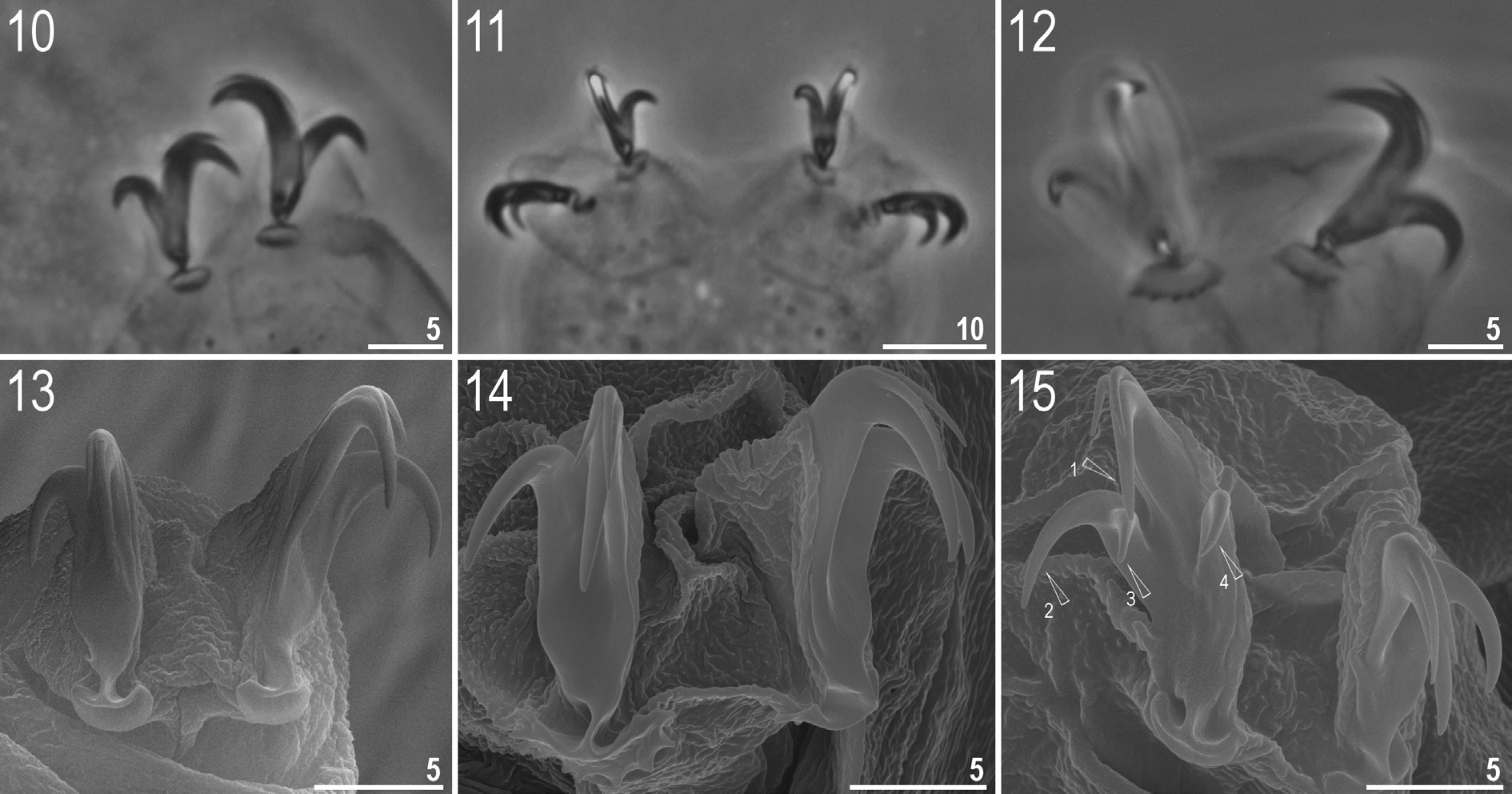
Claws small and slender of the hufelandi type ( Figs 10–14 View Figs 10–15 ). Primary branches with distinct accessory points. Lunules on legs I–III smooth ( Figs 10, 13 View Figs 10–15 ), those on legs IV dentate ( Figs 12, 14 View Figs 10–15 ). Bars under claws absent but paired muscle attachments below claws I–III present (hardly visible under LM and only slightly more visible under SEM).
Eggs (measurements and statistics in Table 4 View Table 4 )
Laid freely, white to light yellow, spherical and with a hufelandi type chorion ornamentation ( Figs 20–28 View Figs 20–23 View Figs 24–29 ). The surface between processes is covered with a dense regular reticulum (mesh diameter 0.5– 0.8 μm) ( Figs 22 View Figs 20–23 , 25–28 View Figs 24–29 ). Processes in the shape of inverted goblets with slightly concave, conical, micro-granulated trunks and well-defined terminal discs ( Figs 22–23 View Figs 20–23 , 25–26, 28 View Figs 24–29 ). When observed under SEM, some process trunks have 3–5 faint, annular ring undulations ( Figs 26, 28 View Figs 24–29 ), whereas in LM these undulations are not visible. Terminal discs are cog-shaped, with each of the 8–10 ‘cog-teeth’ extended to form a long, thin, hair-like and flexible filament that probably serves to enhance the adhesive function of egg processes. Under SEM, small rounded granules or aggregations of granules, 0.06–0.15 μm in diameter, are visible on the filaments ( Fig. 29 View Figs 24–29 ). Central area of the terminal disc with sparse, randomly distributed, small granules ( Figs 25, 28 View Figs 24–29 ). Moreover, the area between the granules on the filaments, the surface of the terminal disk and the trunk of the processes appear, under SEM, to be micro-granulated ( Figs 28–29 View Figs 24–29 ).
DNA sequences
Initially, four molecular markers obtained from four of the eight individuals were sequenced. The sequences from 18S rRNA, 28S rRNA, ITS-2 exhibited a lack of polymorphism, whereas for COI two distinct haplotypes were obtained. Knowing that ITS-2 and COI have relatively high mutation rates, these fragments were sequenced for the remaining four specimens. The eight individuals were found to have identical ITS-2 sequences while COI revealed two haplotypes (frequency 1:1) differing by 20 substitutions. One consensus sequence for each nuclear marker and one consensus sequence for each COI haplotype were deposited in GenBank: 18S rRNA sequence, 1726 bp long (GenBank accession number: KX810008 View Materials ), 28S rRNA sequence, 725 bp long ( KX810009 View Materials ), COI sequence for haplotype 1, 658 bp long ( KX810011 View Materials ), COI haplotype 2, 658 bp long ( KX810012 View Materials ), and ITS-2 sequence, 425 bp long ( KX810010 View Materials ).
Remark
Among the 136 individuals of the new species, one specimen had abnormal claws, i.e., internal and external claw II and III on one leg had additional secondary branches ( Figs 15 View Figs 10–15 , 19 View Figs 16–19 ). Additionally, on the
claws of the IV pair of legs, unique, upwardly turned spurs were present near the lower half of the claw ( Fig. 19 View Figs 16–19 ).
Differential diagnosis
Morphological
Macrobiotus polypiformis sp. nov., by the presence of long, flexible filaments on the egg processes and faint annular undulations on their trunks, is most similar to Ma. paulinae Stec et al., 2015 , but differs from it by possessing larger pores on the cuticle (0.9–1.2 μm in Ma. polypiformis sp. nov. vs 0.3–0.5 μm in Ma. pauliane ), smooth cuticle (seven dorso-lateral patches of sparse granulation arranged symmetrically on both sides of the body in Ma. paulinae ), one patch of granulation on legs I–IV (two distinct patches of granulation, fine and dense granulation above claws and a more robust and sparse granulation in the middle of each leg in Ma. paulinae ), a different number of rows of teeth in the first band of teeth in the oral cavity (3–4 rows in Ma. polypiformis sp. nov. vs a single row in Ma. paulinae ), larger reticulum mesh diameter covering the egg surface (0.5–0.8 μm in Ma. polypiformis sp. nov. vs 0.05–0.2 μm in Ma. paulinae ), trunks of the egg processes with faint annular undulations (distinct and clearly visible in Ma. paulinae ), different morphology of the terminal disc margins (8–10 long, hair-like, flexible filaments in Ma. polypiformis sp. nov. vs small, irregular teeth, instead of filaments, in Ma. paulinae , with only some processes having one to a few flexible filaments), lower number of processes on the egg circumference (19–23 in Ma. polypiformis sp. nov. vs 24–32 in Ma. paulinae ).
Genotypic
It was confirmed, using Basic Local Alignment Search Tool (BLAST; Altschul et al. 1990), that no sequences deposited in GenBank were identical with sequences obtained from the type population of Ma. polypiformis sp. nov. The p-distances calculated for ITS-2 between the new species and Ma. polonicus Pilato, Kaczmarek, Michalczyk & Lisi, 2003 , Ma. spaiens Binda & Pilato, 1984 and Ma. paulinae are 39.4%, 25.6% and 31.1%, respectively. In the case of COI, genetic distances between the two haplotypes of Ma. polypiformis sp. nov. is 3.1%, which is above the threshold for species delineation proposed by Cesari et al. (2009). However, sequences from haplotype 1 and 2 are clearly distant from sequences of each species used in the analysis. For haplotype 1, p-distance range from 20.6% to 25.8%; haplotype 2 from 21.0% to 25.9%. In each case the most similar to the new species is Ma. paulinae ( KT951668 View Materials ) and the most distinct is Ma. macrocalix Bertolani & Rebecchi, 1993 ( HQ876571 View Materials , FJ176208 View Materials -12). Given there is no polymorphism in the ITS-2 sequences coming from the type population and the morphologically most similar species Ma. paulinae differs from the new species by a genetic distance of nearly 21%, it can be claimed that Ma. polypiformis sp. nov. is a valid new species exhibiting two COI haplotypes. P-distances for more conservative DNA fragments (28S rRNA and 18S rRNA) are, as was expected, lower than for more variable markers (COI and ITS-2). The range for 28S rRNA is 7.3%–12.9% with the most similar species Ma. paulinae ( KT935501 View Materials ) and for 18S rRNA 3.0%–6.2% where the closest related species are Ma. paulinae ( KT935502 View Materials ) and Ma. sapiens ( DQ839601 View Materials ).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |