
Rhyacodrilus propiporus, Rodriguez, Pilar & Fend, Steven V., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3664.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C8136C89-7787-477D-BC64-97AA6C16057B |
|
DOI |
https://doi.org/10.5281/zenodo.6160663 |
|
persistent identifier |
https://treatment.plazi.org/id/154FD01D-4B41-FFC4-FF6E-9766BD04CCDB |
|
treatment provided by |
Plazi |
|
scientific name |
Rhyacodrilus propiporus |
| status |
sp. nov. |
Rhyacodrilus propiporus sp. n.
( Figs 9–11 View FIGURE 9 View FIGURE 10 View FIGURE 11 )
Holotype. USNM 1202062, whole-mounted worm stained in hematoxylin and mounted in Canada balsam.
Paratypes. USNM 1202063-65: 1 dissected specimen (14 February 2007), 1 sagittally sectioned (17 March 2007), 1 whole mount (17 March 2007), MNCN 16.03/3081: 1 dissected specimen (17 March 2007), all from the type locality.
Type locality. Beech Swamp, Halifax Co., North Carolina, USA, N36.1435° W77.5551° (17 March 2007).
Other material. 2 dissected and 1 whole mount from Lower Little River at road SR 2031, Harnett Co., N35.2599° W78.8230°, (5 June 2011); 4 dissected, 2 whole mounts, 1 sagittally sectioned (July 2011). 2 whole mounts from Lower Little River at highway NC 217, N35.2637° W78.7406°, (5 August 2011). 6 whole mounts from Lower Little River at highway NC 87, Cumberland Co., N35.4001° W79.1126° (5 August 2011). 2 dissected from Powell's Creek at road SR 1338, Halifax Co., N36.1791° W77.8980° (13 December 2011). 1 whole mount from Friar Swamp at road SR 1740, Columbus Co., N34.3695° W78.4572° (1 March 1998). 1 whole mount from seep at Pettiford Creek, Carteret Co., approx. N34.75° W77.02° (30 January 2009). 1 whole mount from Chinkapin Creek, Hertford Co., N36.2534° W76.8491° (1 March 2001). All material collected by David R. Lenat from North Carolina, USA.
Etymology. From the Latin prope for close and porus for pores. The specific name refers to the very close (nearly median) position of both spermathecal and male pores.
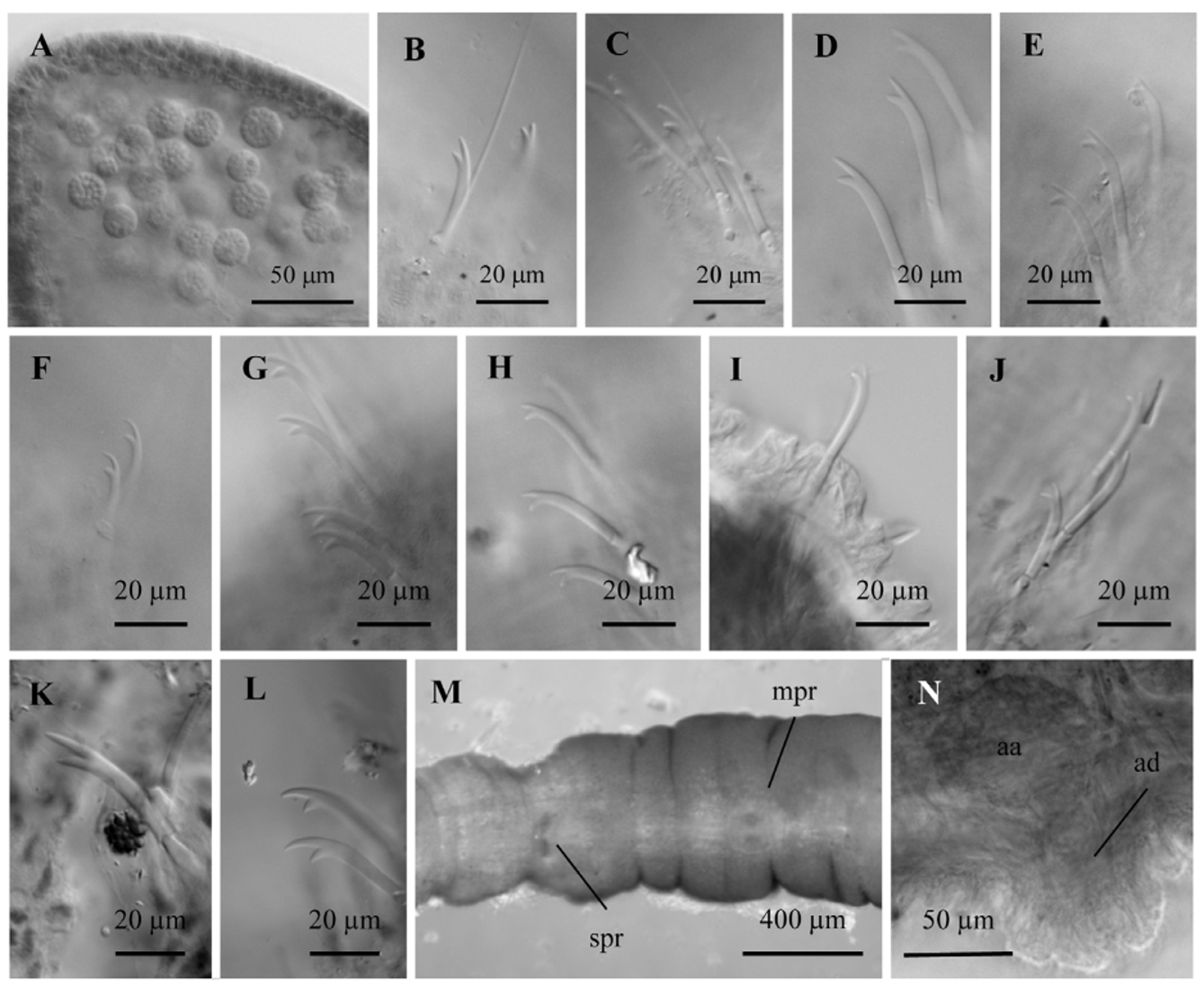
Description (from type locality specimens). Number of segments 63–74. Body diameter 440–456 µm in segment VIII, 556–614 µm in XI. Rounded-conical prostomium 107–202 µm long. Epidermis 7–13 µm high, 25– 45 µm at clitellum. Clitellum from the level of chaetae in X to the end of XII.
Dorsal bundles in anterior segments with 0–3 smooth hair chaetae (229–242 µm long) and 3–4 bifid, nonpectinate chaetae (103–107 µm long) with distal tooth longer than proximal ( Fig. 9 View FIGURE 9 B–E). Hair chaetae absent from segment VIII (or IX) backwards. Posterior dorsal chaetae like ventral chaetae (69–110 µm long). Ventral chaetae bifid; anterior ventral chaetae 4–6 per bundle (73–110 µm long) with distal tooth longer than proximal; posterior ventral chaetae 2–4 (commonly 3) per bundle, 88–99 µm long, and progressively shorter, down to 70–74 µm long in the most posterior segments of complete individuals, distal tooth becoming equal to or shorter than proximal in the most posterior segments ( Fig. 9 View FIGURE 9 F–I).
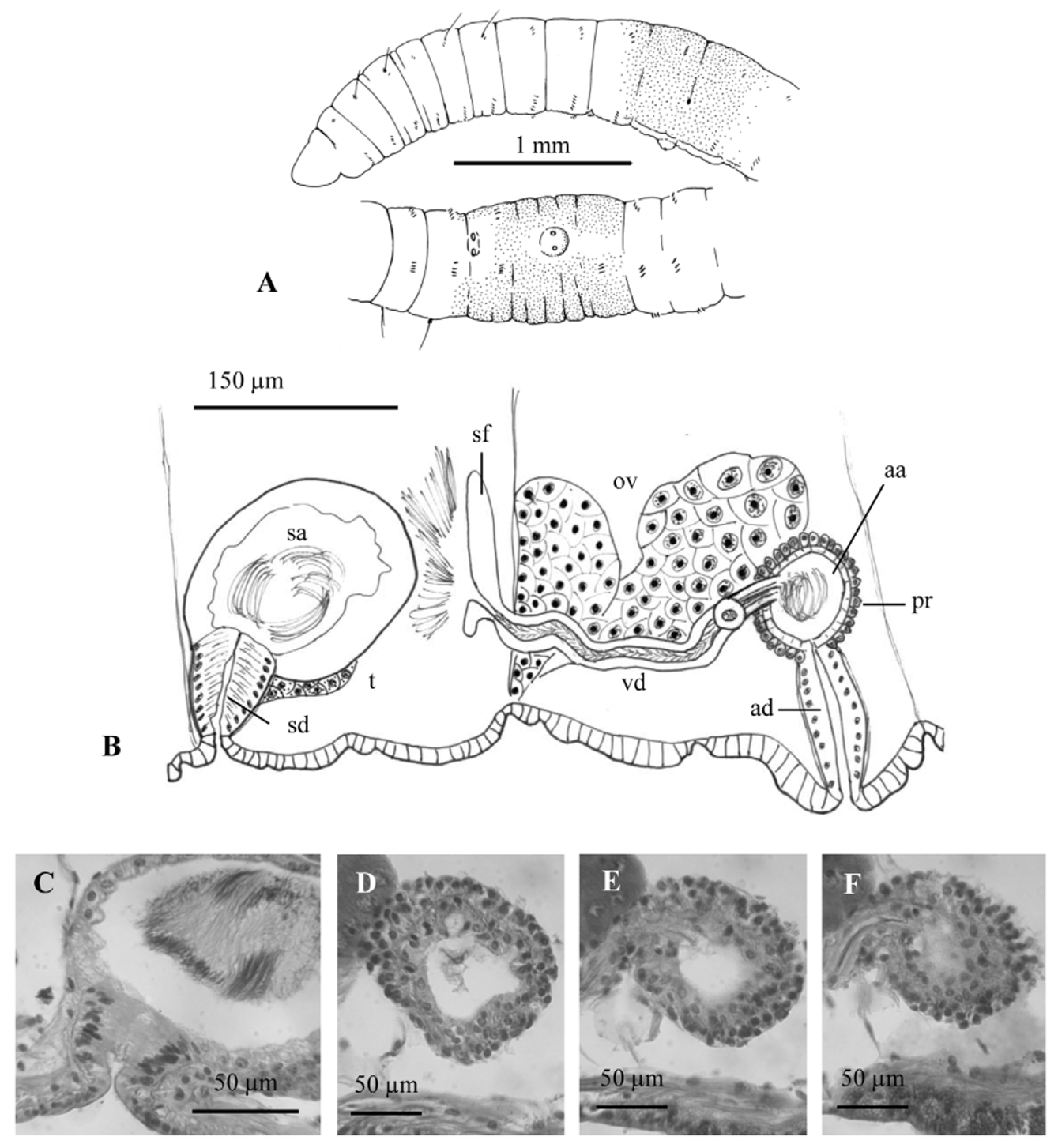
One pair spermathecal pores at the beginning of segment X, close to the mid-ventral line of the body, usually within a shallow, common groove about 150 μm wide; the spermathecal pores surrounded by a thin, muscular ring at the body wall (similar to Fig. 11 View FIGURE 11 C). One pair male pores at about the posterior 1/4 of segment XI, close together near the mid-ventral line, on a prominent porophore 80–120 μm wide and up to 45 μm high, lined with a thin, circular muscle layer ( Figs 9 View FIGURE 9 M, N, 10A). No modified genital chaetae. Ventral chaetae usually absent in X at maturity; in one mated worm a single chaeta was not modified and resembled other ventral chaetae (79 µm long). Female pores in intersegment 11/12, just below lateral line.
Abundant nucleated coelomocytes (diameter 10–24 µm), containing large granules ( Fig. 9 View FIGURE 9 A). Pharyngeal glands in V and poorly developed in IV and VI. Chloragogenous tissue scantily covering the gut from VII backwards, and formed by a thin layer of cells (usually <20 µm high). One pair testes in segment X and one pair ovaries in segment XI. Sperm sac back to segment XV, egg sac back to segment XVI. Spermathecae composed of a short duct (43–77 µm long and 58–64 µm maximum diameter) and an ampulla (154–246 µm long, 138–215 µm wide) in the most anterior part of segment X ( Fig. 10 View FIGURE 10 B, C). Duct lined with columnar epithelium, with a narrow (5– 10 μm) lumen and very thin muscle layer. Ampulla with thin layer of cuboidal epithelium (6–21 μm); lumen with sperm typically arranged in loose clusters.
Male duct consisting of a sperm funnel at the most ventral part of septum 10/11, and a relatively straight vas deferens which joins the atrial ampulla subapically; vas deferens of homogeneous width (diameter 23–24 µm), longer than atrium (ca. 230–315 µm long) ( Fig. 10 View FIGURE 10 B). Atria formed by a spherical ampulla (diameter 57–75 µm) and a duct (78–125 µm long, 43 µm maximum diameter) about 1.5–2 times longer than the ampulla; duct tapering towards the male pore and usually towards the ampulla too. Penis absent. Atrial ampulla with a thin epithelium (7– 8 µm) and very thin musculature, covered by a diffuse layer of prostate cells (up to 10–14 µm high), not forming bundles of cells ( Fig. 10 View FIGURE 10 D–F). The relative size of the atrium is very small, total length less than one third the diameter of the body.
Lower Little River population (supplemental description, differences from type locality population). Body dimensions generally smaller than Beech Swamp population (type locality): number of segments 53–72, diameter 270–400 µm in segment VIII and 360–490 µm in XI (slide-mounted worms). Chaetae similar, but usually shorter than in Beech Swamp population: anterior dorsal bundles with 1–3 smooth hair chaetae (maximum 200–260 µm long, shortest in II and X) and 3–4 bifid chaetae (85–110 µm long) with distal tooth slightly or up to 2 times longer than proximal. Hair chaetae absent from segment X–XII backwards. Posterior dorsal chaetae like ventral chaetae (both 85–100 µm long). Anterior ventral chaetae 4–8 per bundle (95–115 µm long) with distal tooth longer than proximal (sometimes with a thin intermediate tooth). Posterior to clitellum 2–5 ventral chaetae; commonly 3 per bundle in posterior half of worm, with distal tooth becoming equal or shorter than proximal in the most posterior segments.
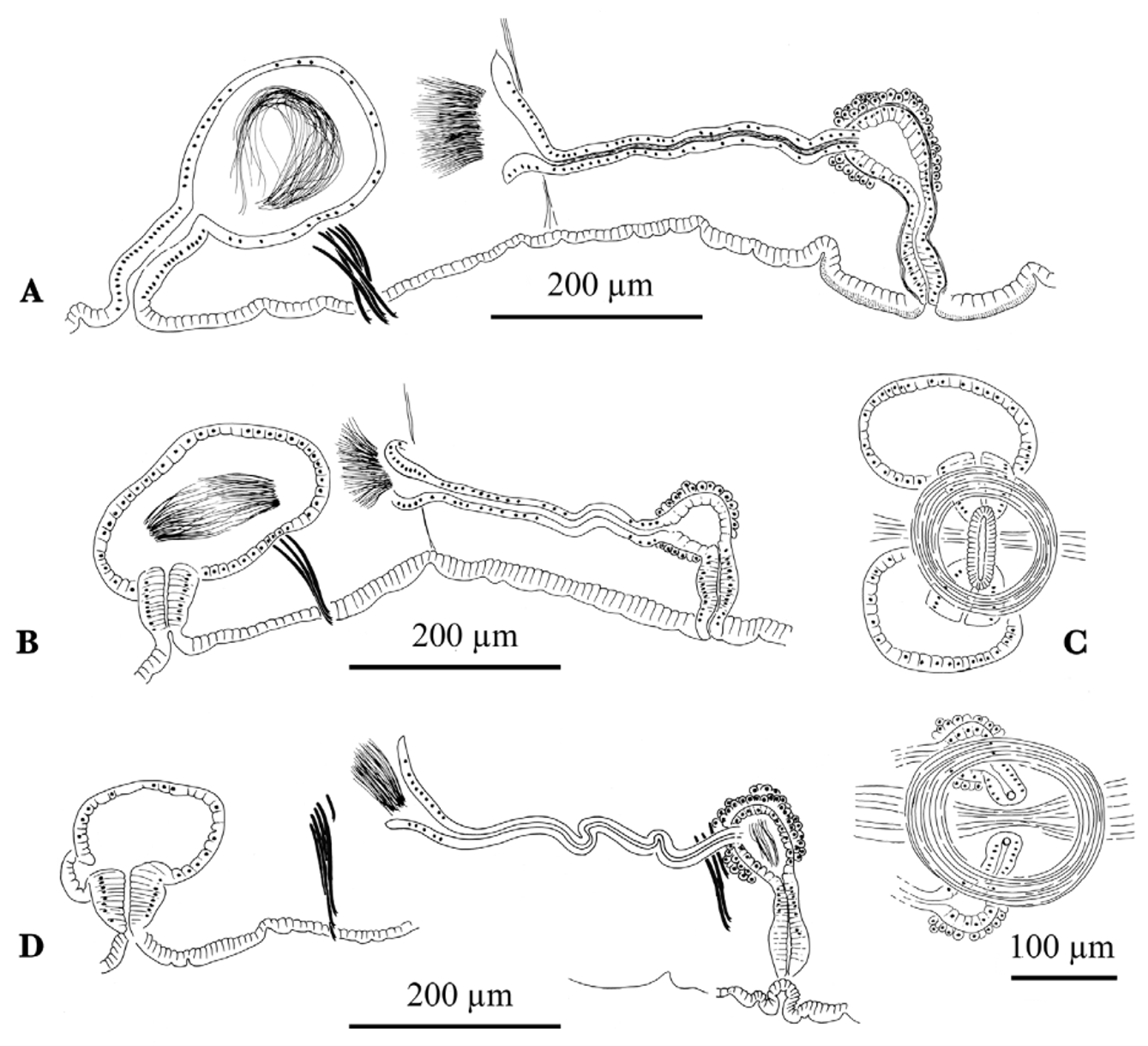
Spermathecal pores usually within a shallow, common groove 50–70 μm wide, pores surrounded by a thin, circular muscle layer ectal to the ventral muscle bands of the body wall ( Fig. 11 View FIGURE 11 C). Male pores on a low porophore about 180–240 μm wide, and up to about 40 μm high; porophore lined with a thin, circular muscle layer.
Spermathecal duct may be cylindrical and up to twice as long as wide, or short and conical, about as wide as long entally ( Fig. 11 View FIGURE 11 A, B) (length 55–96 µm, width 38–62 µm). Spermathecal ampulla ovate (125–245 µm long, 100–180 µm wide in mated worms), usually entirely within X. Atrial ampulla spherical to slightly ovate (60–145 µm wide); duct cylindrical or tapered (55–104 µm long, 30–42 µm maximum diameter); total length of atrium 120–225 µm, about one third to half the diameter of the body at segment XI; relative lengths of duct and ampulla variable, but usually about equal ( Fig. 11 View FIGURE 11 A, B). Atrial ampulla with epithelium 10–20 µm high, very thin musculature, and covered by a diffuse (usually single) layer of prostate cells.
The other listed populations from different locations in North Carolina are represented by limited material, and have been provisionally ascribed to R. propiporus . Specimens from Powell's Creek had very similar reproductive organs, but were unmated and had unmodified ventral chaetae in segment XI ( Fig. 11 View FIGURE 11 D).
Anomalies. One Lower Little River specimen had paired spermathecae in VII and VIII, paired atria in VIII, and a single atrium in IX; only the second male duct had sperm on the funnel. Another specimen had normal reproductive organs, but they were shifted forward by two segments. Six mated specimens had 1 or 2 unmodified chaetae in XI, but these were presumably remnant somatic chaetae, as they were on the line of ventral chaetae, and were not medially directed.
Distribution and habitat. Rhyacodrilus propiporus sp. n. was collected in diverse habitats in eastern North Carolina. Beech Swamp (the type locality) and Powell's Creek are in the Tar River basin, in the northeastern part of the State. Rhyacodrilus habitats were small (2 m wide), sandy streams that typically go dry during summer months. Both streams appear to have good water quality based on the macroinvertebrate composition. Beech Swamp has a diverse oligochaete community, and is the only known location for the monotypic Pilaridrilus Fend and Lenat, 2007 (Lumbriculidae) .
The Little River is in southeastern North Carolina, in the Cape Fear River basin. It is a large (20 m wide), permanent stream, with low gradient and excellent water quality based on the macroinvertebrate community (D.R. Lenat, pers. comm. 2012). Rhyacodrilus propiporus was usually collected from sand and gravel near the middle of the river.
Remarks. Rhyacodrilus propiporus sp. n. belongs to a small group of Rhyacodrilus species with hair chaetae in dorsal bundles and a layer of prostate cells covering the atrial ampulla, and lacking both modified spermathecal and penial chaetae. This group is formed by R. lepnevae Malevich, 1949 emend. Hrabë (1974) ( Russia), and R. lindbergi Hrabë, 1963 a (described from one immature specimen from a cave in Portugal). Martínez-Ansemil et al. (1997) attributed a single specimen with penial chaetae from northwestern Spain to R. lindbergi , although this character was not verified in the type specimen (E. Martínez-Ansemil, pers. comm. 2011). Both species are distinguished externally from R. propiporus sp. n. by the presence of ventral bundles with both simple-pointed and bifid chaetae. Externally, the near-median positions of the spermathecal and male pores distinguish the new species from other Nearctic members of the genus. The new species is also well-separated from other Nearctic Rhyacodrilus by the small size of the atrium and relative lengths of the atrial ampulla and duct (the latter being 1.5 to 2 times the length of the former), and also by the very short, one cell layer prostate covering the ampulla (unique among the Nearctic Rhyacodrilus species).
The population from Lower Little River has been provisionally ascribed to R. propiporus based on similar chaetal characteristics and the peculiar near-median position of male and spermathecal pores. The larger atrium, a relative length of atrial ampulla to duct of about 1, a cylindrical spermathecal duct, and ventral chaetae often present in segment X, distinguish this population from that of the type locality, and its status may be subject to revision with new findings in eastern North America.
This species may have a broader distribution than indicated here. The combination of dorsal hairs limited to anterior segments and anterior dorsal bifid chaetae with the distal tooth longer than the proximal would likely attribute immature R. propiporus to R. subterraneus using the keys by Kathman and Brinkhurst (1998).
Examination of material collected by D. Strayer from several sites in eastern USA, and cited by Kathman and Brinkhurst (1998) as R. subterraneus , revealed no mature specimens, so the presence/absence of penial chaetae cannot be verified. All 6 individuals examined from the USNM collection (# 102832, #187041 and #187042, leg. Strayer), plus 2 additional specimens from the Strayer collection, were immature, with upper tooth of dorsal chaetae thicker and much longer than lower (as in R. subterraneus ), and with short intermediate teeth visible in some individuals but not in others. In the absence of mature worms attributable to R. subterraneus , we conclude that at least some of the North American records of R. subterraneus based on chaetal morphology may be R. propiporus , and that an examination of the male ducts and/or molecular analyses are required to confirm the presence of R. subterraneus in North America (see also remarks for R. clio sp. n. above).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Rhyacodrilinae |
|
Genus |