
Geodia atlantica, (STEPHENS, 1915)
|
publication ID |
https://doi.org/10.1111/zoj.12056 |
|
publication LSID |
lsid:zoobank.org:pub:5CFF222F-0C8D-4FA8-9388-D0C77213710E |
|
persistent identifier |
https://treatment.plazi.org/id/122687EB-FFD0-2161-FC11-CCAE6CE9BA73 |
|
treatment provided by |
Marcus |
|
scientific name |
Geodia atlantica |
| status |
|
GEODIA ATLANTICA ( STEPHENS, 1915) View in CoL
GEODIINAE P ATLANTICA ( PhyloCode SPECIES NAME)
( FIGS 3–6 View Figure 3 View Figure 4 View Figure 5 View Figure 6 , TABLE 1)
Sidonops atlantica, Stephens, 1915: p. 18 View in CoL . Sidonops cf. atlantica, Cárdenas et al., 2010: p. 89 View in CoL .
Geodia sp. , Hougaard et al., 1991a: p. 225, b: p. 470; Warén & Klitgaard, 1991: p. 55; Cedhagen, 1994: p. 67; Klitgaard, 1995: p. 2 (synonymy by this study).
Geodia atlantica, Klitgaard & Tendal, 2004: p. 57 View in CoL ; Todt et al., 2009: table 1; Schöttner et al., 2013: p. 2. Geodia cf. atlantica, Cárdenas et al., 2011 View in CoL : table S1.
Misidentifications:
Isops pallida, Vosmaer, 1882: p. 16 View in CoL (in part).
Geodia macandrewii, Reitner & Hoffmann, 2003 View in CoL : Tafel 2, figure 1.
Geodia barretti, van Soest et al., 2007 View in CoL (in part?): table 2.
Type material examined.
Isops pallida , near Hammerfest, Norway, 71°12′5N, 20°30′5E, 247 m, Willem Barents Exp. 1878–79, RMNH Por 652, wet specimen (only pictures were seen); RMNH, Vosmaer slide collection, box number 37, three spicule preparations with number 64.
Geodia atlantica , holotype, off western Ireland, 54°17′5N, 11°33′5W, 709 m, stones and rock, 9.15 °C, number S. R GoogleMaps .151-27/364-1914.
External morphology and cortex: Colour alive and in ethanol is whitish ( Norway, Flemish Cap) to light brown (Bay of Biscay, Rockall Bank). Choanosome colour alive and in ethanol is brownish (always darker than cortex). Large specimens can be funnel shaped with a deep cavity with irregular swellings and ridges ( Fig. 3A, B View Figure 3 ). From fragments we have seen in large trawl catches at the Faroe Islands, we had the impression that in very large specimens the bottom of the funnel could disappear with time, and that the sponge maintained a ring-shaped wall, 80–100 cm in diameter. The largest specimen found measures 72 ¥ 40 ¥ 39 cm. Other specimens are irregularly plate-shaped and convoluted ( Fig. 3C, D View Figure 3 ). Young specimens are subspherical ( Fig. 3E, G View Figure 3 ). The surface is smooth. The lower sides of the specimens are sometimes covered with stones incorporated in the cortex. Uniporal oscules ( 0.3–0.5 mm in diameter) are scattered on the top surface of small specimens, and oscules are sometimes surrounded by a raised lighter-coloured boundary. Oscules are scattered on the inner side of funnel-shaped specimens so that pores and oscules are on opposite sides ( Fig. 3H View Figure 3 ). Cribriporal pore areas ( 0.3–0.5 mm in diameter) are on the outer side of funnel-shaped specimens and on one side of plate-shaped specimens; they are in small groups, which are evenly scattered over the surface ( Fig. 3I View Figure 3 ). The cortex is elastic, c. 0.5 mm thick (with ectocortex: 30–100 Mm) ( Fig. 4 View Figure 4 ). Anatriaenes within the choanosome are fairly common ( Fig. 4F View Figure 4 ).
Description of type material: Three spicule preparations (with number 64) ( Fig. 4A View Figure 4 ) of the syntype b of Isops pallida . Pictures of the spicules are shown ( Fig. 4B View Figure 4 ). We have seen only pictures of the wet specimen of syntype b: it is a whole spherical specimen about 4 cm in diameter, with very small roots, a thin cortex (< 1 mm thick), slightly raised uniporal oscules and cribriporal pores on opposite sides. It looks similar to specimen ZMAPOR 21406a from Norway ( Fig. 3F View Figure 3 ).
The holotype of G. atlantica is a small subspherical specimen (2.7 ¥ 2 cm). This specimen is represented in plate II of Stephens (1915) and Figure 3G View Figure 3 . In the Dublin Museum , there are also five spicule preparations made by Stephens (four spicule slides and one section). New thick sections were made for this study ( Fig. 4C, D View Figure 4 ). Figure 5 View Figure 5 shows SEM pictures of this holotype .
Spicules ( Figs 4–5 View Figure 4 View Figure 5 , Table 1): Megascleres: (a) oxeas, straight or bent, length: 1275–4440 Mm; width: 11–68 Mm. (b) Orthotriaenes, rarely dichotriaenes, straight or slightly bent rhabdome, rhabdome length: 630–4400 Mm; width: 18–125 Mm; orthotriaene clad length: 95–750 Mm; protoclad length: 190–430 Mm; deuteroclad length: 90–300 Mm. (c) Anatriaenes, straight or slightly bent rhabdome, rhabdome length: 376–5200 Mm; width: 2–32 Mm; clad length: 9–300 Mm. (d) Protriaenes, very rare [one reported in the type ( Stephens, 1915) and one observed in PC626], rhabdome length: 3000 Mm; width: 8–15 Mm; clad length: 96–130 Mm. Microscleres: (e) sterrasters, slightly elongated, more rarely spherical, length: 80–125 Mm; width: 75–112 Mm; thickness: 70–88 Mm. Rosettes are made of 2–6 smooth rays; rosette diameter: 4–7 Mm; hilum diameter: 10–20 Mm. (f) Spheroxyasters, rough actines, 5–16 Mm in diameter. (g) Oxyasters I, 3–8 rough actines, diameter: 22–110 Mm [maximum measured in type by Stephens, (1915)]. (h) Oxyasters II, 9–25 rough actines, usually with a larger centrum than oxyasters I, diameter: 12–35 Mm.
DNA barcodes: GenBank accession nos. HM 592679 View Materials , HM 592695 View Materials , EU442195 View Materials ( Folmer COI): we have sequenced specimens from western and northern Norway (10) , Rockall Bank (1), and Flemish Cap (1): the Folmer COI is identical in all these specimens. No. KC481227 View Materials (18S), obtained from ZMBN 77927 View Materials ( Korsfjord , Norway) .
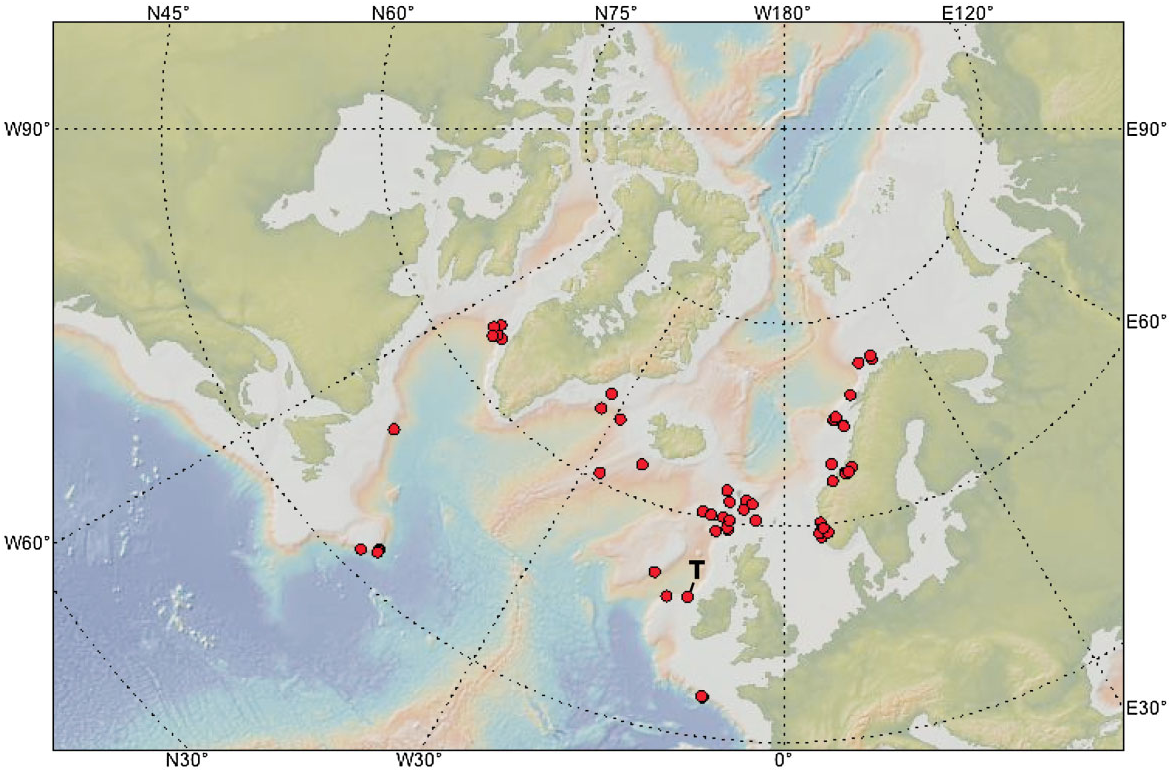
Distribution ( Fig. 6 View Figure 6 ): This species has an amphi- Atlantic boreal distribution. It has been recorded at depths of 65–2338 m, the shallowest record being from divers in Sandsfjord, Rogaland, Norway ( Moen & Svensen, 2008), the deepest boreal records being from south of Iceland and off south-east Greenland, while the deepest record overall is from the Bay of Biscay. Geodia atlantica seems only to be present in the south-western Barents Sea and absent in Arctic waters, which might explain why it is not mentioned by Koltun (1966). It has been found at temperatures between 1.4 °C ( Denmark Strait) and 10.5 °C, but is usually found at temperatures higher than 3 °C.
Biology: We found no indications of asexual reproduction. The predatory chiton Hanleya nagelfar Lovén, 1846 and the parasitic foraminiferan Hyrrokkin sarcophaga Cedhagen, 1994 have been found living on G. atlantica ( Warén & Klitgaard, 1991; Cedhagen, 1994; Todt et al., 2009). Sea urchins are also possibly feeding on this sponge; the two species observed in Figure 3C View Figure 3 were tentatively identified from the photo as Cidaris cidaris (L., 1758) and Gracilechinus alexandri ( Danielsen & Koren, 1883) (T. Saucède, pers. comm.). Cidaris cidaris ( Rouho, 1888; Mortensen, 1928) and other cidarids ( Bo et al., 2012) are indeed considered to be sponge predators whereas G. alexandri is more of an omnivore opportunist which may be more interested in the small organisms living around and on the sponge. Other associated fauna has been investigated by Klitgaard (1995). The chemistry (elemental analysis, amino acids, sterols, and quaternary ammonium compounds) has been investigated by Hougaard et al. (1991a, b).
Distinctive characters: External morphology: The deep funnel shape or plate convoluted shape, with smooth surface. The pattern of distribution of pores and oscules: when one finds a fragment of a funnel or plate-shaped specimen, oscules are on one side, pores on the other. Spicules: Lack of microxeas (as in G. phlegraei and G. parva ) and very common anatriaenes.
Remarks: Burton (1930) synonymized I. pallida with G. phlegraei by stating that he had compared type
Means are in bold; other values are ranges; N = 30 unless stated otherwise in parentheses, or unless measurements come from other studies. A dash indicates that this measurement is not given in the literature. n.f., not found; n.o., not observed in the subsample in our possession.
slides from the ‘Norman collection’, and taxonomists followed his conclusions. Even Vosmaer (1933: 141–142) accepted the synonymy after having examined a slide of G. phlegraei sent to him by Sollas. However, the only slides of I. pallida that we found in the Norman collection (BMNH 10.1.1.1149 to 1156 and MNHN-DN45) had labels saying ‘ Isops pallida Vosmaer /Lervig, Norway, 1879’. In 1879, Norman did stay in Leirvik (current name of ‘Lervig’) on the island of Stord in the Hardangerfjord in western Norway ( Norman, 1893), so these are clearly not from the type of I. pallida (which was collected near Hammerfest in northern Norway). Interestingly, the thin cortex ( 0.5 mm), the large spiny oxyasters, and spheroxyasters showed that these slides from the Norman Collection were not from a G. phlegraei but from a G. atlantica . So Burton (1930) had probably not examined type slides and had not noticed the difference between G. atlantica and G. phlegraei spicules. However, in the course of this study, Rob van Soest (Naturalis, Leiden) rediscovered in the Vosmaer slide collection (box 37) five slides with labels stating ‘ Isops pallida Vosm’ and ‘Bar. I p. 16’ ( Fig. 4A View Figure 4 ) which was understood as a reference to the Barents Sea expedition 1st publication, and to the page number of the original description of I. pallida in Vosmaer (1882). We therefore concluded that these five slides were the syntype slides of I. pallida . Three slides had the number 64 while two slides had the number 62 ( Fig. 4A View Figure 4 ); the spicule morphologies and abundance corresponded to the descriptions given of syntype a (= 62) and syntype b (= 64), the latter being the one represented in the original plates (external morphology and spicules). Indeed, Vosmaer (1882) clearly notes that specimen a has considerably fewer oxyasters and anatriaenes than specimen b. This is because Vosmaer (1882) simply mixed one specimen of G. phlegraei (a) with one specimen of G. atlantica (b): spicule morphologies (spheroxyasters, large spiny oxyasters, abundant anatriaenes) ( Fig. 4B View Figure 4 ) and sizes ( Table 1) clearly show that specimen b of I. pallida type material is conspecific with G. atlantica . Since Vosmaer, (1882) did not explicitly designate a holotype, we have the possibility to designate a lectotype. We formally designate the syntype a as the lectotype of Isops pallida Vosmaer, 1882 . Recommendation 74B of the ‘International Code of Zoological Nomenclature’ states that in choosing a lectotype among syntypes, preference should be given to the illustrated specimen, in our case syntype b. But we decide to go against this recommendation for the following reasons: (1) a comes before b in the alphabet, (2) atlantica has been used far more in the literature than pallida and, above all, (3) I. pallida has always been considered a junior synonym of G. phlegraei (so our deci- sion will preserve the stability of the nomenclature). Following our decision, specimen b then becomes a misidentification, and I. pallida does not become a junior synonym of G. atlantica .
Just before the publication of this revision, Rob van Soest discovered in 2013 a jar (RMNH Por. 652) labelled: ‘ Isops sphaeroides Vosm (type v. I. pallida Vosm. ) W. Barents exp. 1878/79, 71°12′5″N 20°30′5″O, Coll. G.C.J. Vosmaer 12 Juli 1879 ’. Inside are two specimens and another small label written in pencil ‘ Isops pallida, N. Archive Suppl. 1’ which refers to the original description of I. pallida by Vosmaer in ‘Niederlaendisches Archiv fuer Zoologie Supplementband 1’. We therefore believe that these two specimens are the two syntypes of I. pallida . The external morphologies (observed from pictures, courtesy of R. van Soest) of the largest specimen (a = lectotype) and of the smallest specimen (b) confirm the above conclusion based on spicules: a is a G. phlegraei and b a G. atlantica .
We noted that G. atlantica had a second smaller category of oxyasters that Vosmaer (1882) has seen in specimen b, but not Stephens (1915). Stephens (1915) states that the cortical spheroxyasters become larger in the choanosome; these are actually the oxyasters II. Admittedly, spheroxyasters and oxyasters II can be difficult to separate in spicule preparations, unless carefully measured and examined in thick sections (spheroxyasters are in the ectocortex, oxyasters II usually in the choanosome just below the cortex). In some specimens anatriaenes may be separated into two size categories (e.g. UPSZMC 78293 from the Flemish Cap), especially based on the rhabdome length (376–530 vs.> 2000 Mm), but since a continuum of anatriaene sizes exists in other specimens (e.g. PC222 from northern Norway), we refrained from doing so. There is usually a clear predominance of orthotriaenes over dichotriaenes (but not always the case, see MNHN-ThalassaZ407).
The main difference between the type and the Norwegian specimens is that, in the Norwegian specimens, the asters are less spiny, and the oxyasters I are smaller and much less abundant ( Fig. 5 View Figure 5 ); this may be due to the shallower environment of the Norwegian specimens ( 200–400 m) compared with the type ( 709 m). No consistent morphological differences were found between specimens from the Flemish Cap and specimens from the NEA. A more NEA southern morph may be present (found in the MNHN Thalassa and Centob collections). These specimens are irregularly plate shaped, with a darker external colour and are usually found growing around coral. We have never seen it with the characteristic funnel shape but we have only seen small specimens (less than 15 cm long) .



| RMNH |
National Museum of Natural History, Naturalis |
| R |
Departamento de Geologia, Universidad de Chile |
| HM |
Hastings Museum |
| COI |
University of Coimbra Botany Department |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Geodia atlantica
| Cárdenas, Paco, Rapp, Hans Tore, Klitgaard, Anne Birgitte, Best, Megan, Thollesson, Mikael & Tendal, Ole Secher 2013 |
Geodia atlantica
| Schottner S & Hoffmann F & Cardenas P & Rapp HT & Boetius A & Ramette A 2013: 2 |
| Klitgaard AB & Tendal OS 2004: 57 |
Geodia sp.
| Klitgaard AB 1995: 2 |
| Cedhagen T 1994: 67 |
| Hougaard L & Anthoni U & Christophersen C & Larsen C & Nielsen PH 1991: 225 |
| Waren A & Klitgaard A 1991: 55 |
Sidonops atlantica , Stephens, 1915 : p. 18
| Cardenas P & Rapp HT & Schander C & Tendal OS 2010: 89 |
| Stephens J 1915: 18 |
Isops pallida , Vosmaer, 1882 : p. 16
| Vosmaer GCJ 1882: 16 |