
Tyrannosaurus rex, Osborn, 1905
|
publication ID |
https://doi.org/10.1002/ar.24118 |
|
DOI |
https://doi.org/10.5281/zenodo.4582709 |
|
persistent identifier |
https://treatment.plazi.org/id/112887D5-FFA7-FF90-FF56-FB76FEFFFA0B |
|
treatment provided by |
Jeremy |
|
scientific name |
Tyrannosaurus rex |
| status |
|
Henry Fairfield Osborn first introduced Tyrannosaurus rex to the scientific community and the public in 1905 ( Osborn, 1905). It has reigned as the quintessential giant carnivorous dinosaur ever since ( Hone, 2016). However, as studies of T. rex and the rest of the Tyrannosauridae have progressed, it has become increasingly apparent that T. rex is far from typical and representative of the theropod lineage as a whole. Tyrannosaurids constitute a highly derived and anatomically unusual group, possessing wide and deep skulls, stout teeth, elongate hindlimbs with arctometatarsalian pedes, and reduced forelimbs with only two functional digits ( Brusatte et al., 2010). Phylogenetically, tyrannosaurids are members of the Coelurosauria, which otherwise consists of predominantly small-bodied theropods. Tyrannosaurus rex stands out in representing the terminal extreme for gigantism within Tyrannosauridae ( Erickson et al., 2004; Holtz, 2004). The iconic size of T. rex has led to myriad questions pertaining to its development ( Erickson et al., 2004; Erickson and Currie, 2006), how its great size evolved ( Erickson et al., 2004), and the ecological and physical upper bounds of terrestrial predator existence ( Alexander, 1985; Henderson and Snively, 2004; Holtz, 2008). Nevertheless, since T. rex ’s discovery other enormous carnivorous dinosaurs belonging to the non-coelurosaurian tetanuran lineages Allosauroidea and Spinosauridae have been found that are purported to be nearly as large as T. rex , if not larger ( Currie, 1996; Dal Sasso et al., 2005), suggesting T. rex’ s giantism does not stand out among nonavian theropods.
ABBREVIATIONS: AMNH = American Museum of Natural History, New York, New York; BHI = Black Hills Institute of Geological Research, Hill City, South Dakota; CM = Carnegie Museum of Natural History, Pittsburgh, Pennsylvania; FMNH = Field Museum of Natural History, Chicago, Illinois; FSAC = Faculté des Sciences Aïn Chock, Casablanca, Morocco; MOR = Museum of the Rockies, Bozeman, Montana; MSNM = Museo di Storia Naturale di Milano, Milano, Italy; MUCPv-CH = Museo de la Universidad Nacional del Comahue, El Chocón collection, Neuquén, Argentina; RSM = Royal Saskatchewan Museum, Regina, Saskatchewan, Canada; TMP = Royal Tyrrell Museum of Palaeontology, Drumheller, Alberta, Canada
RSM P2523.8 ( RSM: Royal Saskatchewan Museum, Regina, SK, Canada) was discovered in an iron-stained, heavily cemented sandstone lens of the Frenchman Formation (Maastrichtian, Upper Cretaceous) near Eastend , Saskatchewan, Canada in 1991. However, excavation did not begin in earnest until 1994. The extremely hard matrix surrounding the fossils, in combination with the size of the specimen, led to a prolonged period of preparation and delays in disseminating its discovery and importance regarding theropod giantism. Although some measurements of the limb bones have been disseminated in previous studies ( Larson, 2008a; Benson et al., 2014; Campione et al., 2014), including inaccurate values (owing to being made prior to full preparation of the specimen]), RSM P2523.8 has never been formally described and its skeletal proportions scientifi- cally quantified.
DESCRIPTION
Overview
RSM P2523.8 is represented by an associated but disarticulated skeleton ( Fig. 1 View Fig. 1 ). Of special note is the abundance of incompletely coossified elements that were previously used as indices of somatic immaturity for the taxon ( Currie et al., 2003). These include quadrates that are not fused to the quadratojugals, surangulars that are not fused to the articulars, and prominently visible open sutures among the basisphenoid, exoccipital-opisthotic, and prootic elements. Some fusions within RSM P2523.8 ’s skull are also not bilaterally symmetrical. For instance, the right jugal osteoderm [the rugose cheek horn common to many tyrannosaurs ( Currie et al., 2003)] is fused, with the suture fully obliterated. Nevertheless, its left counterpart is completely unfused ( Fig. 1 View Fig. 1 ). The fourth sacral centrum is not fused to either the third or the fifth sacral centra. Numerous vertebral neural arches are not fused to the centra ( Fig. 2 View Fig. 2 ), and the scapula is not fused to the coracoid ( Fig. 3 View Fig. 3 ).
As is common among tyrannosaurs ( Brusatte et al., 2010), RSM P2523.8 is riddled with skeletal pathologies. These include rugosities and deformations surrounding the neural canals of caudal vertebrae 15–17, a thick ingrowth of interdental bone within the eighth alveolus of the left maxilla ( Fig. 4 View Fig. 4 ) and an incompletely healed and fractured dorsal rib head from the right side of the animal ( Fig. 5 View Fig. 5 ).
Diagnosis
RSM P2523.8 is referred to T. rex based on the following suite of anatomical characteristics: (1) premaxillary teeth arranged in a laterally oriented tooth row and with D-shaped cross-sections; (2) a maxilla with a rounded rostral margin and a convex ventral margin; (3) “keyhole-shaped” and anteriorly directed orbits; (4) a corneal boss posterodorsal to the orbit; (5) a broad and elongate quadratojugal process and pneumatic excavation of the squamosal; (6) a maximal skull width estimated to be nearly two-thirds that of the premaxilla-occipital condyle length; (7) a long and strap-like scapula with accentuated proximal expansion; (8) a pubic shaft showing concave anterior curvature; and finally (9) an anteriorly projecting sheet of bone spanning roughly half the dorsoventral length of the ischium.
Histology
Transverse petrographic microscopy analysis reveals that the diaphysis of the right fibula to be almost entirely composed of dense Haversian bone ( Fig. 6A,B View Fig. 6 ). Between two and five tightly spaced lines of arrested growth, collectively constituting an External Fundamental System ( Cormack, 1987), are locally present nearest the lateral periosteal surface of the element ( Fig. 6A View Fig. 6 ).
Our histological sampling of the fibula of RSM P2523.8 reveals porous, poorly organized bone matrix composing the cortex spanning from the medial periosteal surface to approximately 20% of the centroid ( Fig. 6C View Fig. 6 ). There is no definitive evidence of medullary bone associated with ovulation ( Schweitzer et al., 2005). Although reminiscent of previous descriptions of medullary bone in T. rex , the inner region of the element shows extensive bioerosion in the form of radiating Wedl tunnels ( Trueman and Martill, 2002) ( Fig. 6D View Fig. 6 ). The diagenetic activity appears to have remodeled the former secondary osteons, whose partial borders are evident from the histology, leading to unorganized mineral globules with only superficial similarities with medullar bone structure ( Schweitzer et al., 2005).
Skull Elements
The right premaxilla is incomplete ( Fig. 7 View Fig. 7 ), but its total size is estimated at approximately 140 mm in anteroposterior length (based on comparison with BHI 3033 ). The premaxilla includes the symphysis and the outer walls of four alveoli.
In lateral view, the right maxilla of RSM P2523.8 has a strongly convex alveolar margin ( Fig. 8 View Fig. 8 ). The maxillary fenestra spans 122 mm at its greatest anteroposterior length, while the antorbital fenestra spans 280 mm. Ten of the presumed 11 alveoli are preserved. On the ventral surface of the maxillary shelf, there are pits above dentary tooth positions 2–10. As in other tyrannosaurids, other than Albertosaurus sarcophagus ( Currie, 2003) , only the first four pits are pronounced.
The nasals are fused and are nearly complete, lacking only the slender tips of the narial processes ( Fig. 9 View Fig. 9 ). The nasals are 790 mm long, with the missing tips likely adding an additional 10 mm (based on comparison with BHI 3033). The external surfaces of the nasals are heavily sculptured. The contact with the maxilla is asymmetrical and is not fused. However, independent movement of either bone would have been restricted by interfingering staircase sutures, as is the case in large specimens of Daspletosaurus torosus , Tarbosaurus bataar , and other specimens of T. rex ( Hurum and Sabath, 2003).
As in other tyrannosaurids, a shallow concavity on the medial surface of the jugal has an opening that connects with the jugal sinus ( Fig. 10 View Fig. 10 ). There is a large circular pneumatic fenestra in the posteroventral corner of the antorbital fossa. A large rugosity is present on the posteroventral corner of the left jugal. This rugosity, which indicates the presence of a blunt jugal “horn,” is absent on the right jugal. It is known from other tyrannosaur specimens that this rugosity is a separate ossification that typically becomes fused to the jugal during ontogeny.
The left lacrimal is nearly complete ( Fig. 11 View Fig. 11 ). It measures 360 mm in anteroposterior length and 375 mm in dorsoventral height. The two arms meet at an acute angle of slightly less than 90 degrees.
Supraorbital ossifications are fused to rugosities on both postorbitals ( Fig. 12 View Fig. 12 ). The postorbital boss is 6 cm long, 14 cm high, and 7 cm thick (lateromedial).
The right squamosal is exceptionally well preserved ( Fig. 13 View Fig. 13 ). There are two large pneumatophores in the posteromedial half of the medial pneumatic chamber. The larger of these pneumatophores (measuring 78 mm in dorsoventral height and 55 mm in anteroposterior length) appear in the region dorsal to the quadrate cotyle. A smaller anteroventral pneumatophore (measuring 18 mm in dorsoventral height and 35 mm in anteroposterior length) invades the medial anteroventral portion of the squamosal.
The quadrates are tall (386 mm in dorsoventral height) ( Fig. 14 View Fig. 14 ), but are not fused to the quadratojugals. This is atypical for mature tyrannosaurids. Relative to the anteroposterior length of the quadrate wing of the pterygoid, the pterygoid wing of the quadrate is unusually long (22 cm) for a tyrannosaurid. Ventromedial to the pterygoid process, a 50 × 30 mm pneumatic sinus enters the anterior surface. The anterolateral–posteromedial axis of the articular condyles for the lower jaw is 18 cm wide. The quadrate–quadratojugal fenestra is large with a dorsoventral diameter of roughly 10 cm.
The quadratojugal of RSM P2523.8 extends approximately 15 cm into the lateral temporal fenestra ( Fig. 15 View Fig. 15 ). However, the dorsal edge of the quadratojugal shows only a shallowly overlapping contact with the long quadratojugal process of the squamosal.
The braincase is nearly complete and has suffered only slight taphonomic distortion ( Fig. 16 View Fig. 16 ) with the cultriform process and most of the left paroccipital process being the only notable absences. The parietals and frontals are tightly bound to each other and to the rest of the braincase by complex interdigitating sutures. However, these are not fused. The orbitosphenoid, septosphenoid, sphenethmoid, and part of the orbital septum are fused. The frontals are partially fused, and are dorsoventrally deep between the supratemporal fossae and interorbital region. As a result, the contacts with the nasals and prefrontals slope anteroventrally. The lacrimal suture is 11 cm deep dorsoventrally and the postorbital suture is 9 cm deep dorsoventrally. The fused parietals measure 195 mm across the midline, 270 mm between the laterosphenoid contacts, and roughly 600 mm between the distal ends of the suture boundary between the paroccipital processes. The nuchal crest is incomplete on the left side, but was clearly asymmetrical, being thinner on the right side (37 mm anteroposteriorly compared with 43 mm). The parietals form a pronounced sagittal crest. The laterosphenoid extends dorsolaterally into the postorbital process roughly 21 cm from the midline and more than 5 cm beyond its contact with the frontal and parietal. The dorsal surface of the postorbital process of the laterosphenoid is smooth where it contacts a trough-like surface in the postorbital. The supraoccipital is fused indistinguishably onto the occipital surface of the parietal, as is the case in the other T. rex specimens— TMP 1981.6.1 , FMNH PR2081 and MOR 555 — but unlike other tyrannosaurids, which tend to have pronounced knob-like processes that extend conspicuously posterior to the parietal. The basipterygoid process is 10 cm below the level of the basal tuber. The basipterygoid processes are 24 cm across. The basioccipital and exoccipitals are indistinguishably fused in the occipital condyle. The prootic is fused anteriorly with the basisphenoid and laterosphenoid, and the line of contact cannot be seen. However, the exoccipital-opisthotic, prootic, and basisphenoid sutures are visible.
The left ectopterygoid is better preserved than the right ( Fig. 17 View Fig. 17 ), and is inflated to a thickness of more than 7 cm. A large, 73 × 31 mm, medially oriented pneumatic opening leads into a sinus that hollows out the bone and extends into at least the base of the jugal process. There also seems to be a small, 18 × 9 mm pneumatophore in the base of the hooked jugal process.
The minimum length of the right dentary of RSM P2523.8 is 810 mm, but the posterior edge is missing ( Fig. 18 View Fig. 18 ). At the level of the sixth alveolus, the left dentary is 94.5 mm wide mediolaterally. The tooth row consists of 14 alveoli on each side and is 595 mm long. The mylohyal foramen of the splenial has a dorsoventral diameter of 5.5 cm.
The surangulars are 645 mm long, with maximum depths of 250 mm, but in spite of their great sizes, neither is fused with the articular ( Fig. 19 View Fig. 19 ). The adductor shelf protrudes 4 cm over the lateral surface. As in most other tyrannosaurids, the main vertical body of the surangular is pierced by sporadic openings. The insertion on the articular for the depressor mandibular musculature is 20 cm across and faces more posteriorly than dorsally. Posteromedial to the glenoid is a large pneumatophore that is 25 mm in diameter.
SIZE ESTIMATION
To quantify the size of RSM P2523.8 and compare it to that of other T. rex specimens ( n = 11), a series of skeletal measurements were taken. To help ensure consistency of measurement technique, all measurements on all specimens were made by one of the authors (P.J.C.). All measurements were made by hand using standard digital calipers or measurement tape, as the size of the measured element dictated ( Table 1).
The standard in neontology for contrasting size across animals of disparate proportions is to use body mass ( Campione et al., 2014; Erickson, 2014). Campione et al. (2014) offered a revised equation for using minimal femoral circumference (MFC) to estimate total body mass across nonavian bipeds, dinosaurs in particular (log 10 M = 2.754 · log 10 C − 0.683, where M is body mass and C is minimum femoral circumference). One limitation of such approaches is that the hollow femoral shafts of theropods are prone to taphonomic distortion. The mid-femur shaft of RSM P2523.8 is slightly damaged, but its minimum transverse diameter (199 mm) is not deformed and indicates a femoral circumference of approximately 590 mm (based on the ELLR method of tyrannosaur femoral circumference estimation, Bradley et al., 2015). The body mass of RSM P2523.8 is thus estimated to be ~8,870 kg ( ± 25%).
Following from the 590 mm femoral circumference estimation, the relative femoral section moduli ( Erickson et al., 2002; Therrien and Henderson, 2007) of RSM P2523.8 is calculated at 650,289 cm 3 and that of FMNH PR2081 , the next largest T. rex specimen, is 617,781 cm 3. Section moduli therefore suggest that the RSM P2523.8 femur had a little more than 1.05 times the bending strength of FMNH PR2081 .
Snively (2018) suggested that comparing the surface area of the last presacral vertebrae offers another potential method of assessing relative body mass between large theropod specimens of the same taxon. Assuming a roughly horizontal body position, the presacral vertebral column of a theropod dinosaur is analogous to a cantilevered-beam loaded in bending. Gravity exerts a torque anterior to a given vertebra, equal to the distance from the vertebra to the center of mass anterior to it multiplied by the mass of this portion of the body. A vertebral centrum is subject to shear stress from this bending moment. The ability of a vertebral centrum to withstand these forces is likely proportional to the surface area of the anterior centrum face (just as the ability of a femoral shaft to withstand compressive forces is proportional to its cross-sectional area). As measured using the imaging software imageJ ( Schneider et al., 2012), the surface areas of the last presacral anterior centra face in RSM P2523.8 and FMNH PR2081 are 650 and 625 cm 2, respectively. This suggests that the presacral loadings of RSM P2523.8 were 1.04 times greater than those of FMNH PR2081 , very near the discrepancy in bending strength calculated based on femoral crosssectional area.
Size comparisons of RSM P2523.8 with the largest specimens of other giant theropod genera were also made using the femoral circumference method of Campione et al. (2014) ( Table 2 View TABLE 2 ).
The Challenge of Sizing Spinosaurus aegyptiacus
Although using MFC to estimate bipedal dinosaur body mass has emerged as a leading technique and has been quantitatively calibrated and evaluated through comparisons with modern animals ( Campione et al., 2014), it should be remembered that the approach makes several assumptions about the biology of the taxa it is applied to. The theropod S. aegyptiacus presents a particular challenge for this technique, because the locomotion and habits of these taxa are thought by many to fundamentally differ from that of most other theropods.
The S. aegyptiacus mass estimation here presented should be interpreted with great caution, as many lines of evidence suggest it may be a substantial underestimation. This estimation was arrived at by using the reconstruction of S. aegyptiacus created by Ibrahim et al. (2014), which isometrically scaled know material, including the femur of specimen FSAC-KK 11888, to the size of MSNM V4047—a specimen that includes only a skull. Whether or not isometric scaling is reasonable to assume across intraspecific theropod specimens of great size variability is a debatable issue. The MFC method assumes that MFC relates directly to minimal femoral cross-sectional area, and, therefore, to the strength of the femur. Ibrahim et al. (2014) reported on the internal femoral shaft structure of S. aegyptiacus , and found it to be unique among theropods in lacking an open medullary cavity. This may have made the cross-sectional strength of S. aegyptiacus femora proportionally greater, relative to femoral circumference. Various researchers have also postulated that S. aegyptiacus was partially quadrupedal or even obligately quadrupedal ( Ibrahim et al., 2014), again unlike all other theropods. If so, the femur of S. aegyptiacus would not have been the sole weight bearer and a proportionally greater body mass could be expected relative to femoral circumference. Finally, Ibrahim et al. (2014) suggested that S. aegyptiacus was adapted for a semiaquatic lifestyle, with greatly reduced hind limb proportions. Being semiaquatic, S. aegyptiacus may have frequently relied on buoyancy in water to support its weight, and, if so, its femoral proportions may be misleading.
Furthermore, if Ibrahim et al.’s Ibrahim et al. (2014) interpretation is correct and the hindlimbs of S. aegyptiacus were reduced in all dimensions relative to its body mass, mass calculations based on MFC would not be expected to yield accurate estimations. As such, it may not, at present, be possible to reasonably estimate the mass of S. aegyptiacus . See also Therrien and Henderson (2007)) for a further discussion of the challenge of estimating the body mass of MSNM V4047.
Comparisons with Other T. rex Specimens
Substantial variation in skeletal proportions has been previously recognized among adult specimens of T. rex , and, although strong statistical support is lacking ( Mallon, 2017), some researches have grouped T. rex specimens into two morphotypes: a gracile form, with more elongate and slender skeletons, and a robust form with stockier proportions and greater estimated mass relative to body length and height ( Larson and Frey, 1992; Larson, 2008b). This purported dichotomy is contentious, with arguments over whether the differences reflect sexual dimorphism or are simply the manifestation of a gradational ontogenetic continuum and/or individual variation ( Carpenter, 1990; Larson and Frey, 1992; Larson, 1994; Larson, 2008b). Those in the former camp speculate that T. rex shows reversed sexual dimorphism ( Larson and Frey, 1992; Larson, 2008b). As in many reptiles and birds, it is suggested that T. rex females displayed a more robust morphotype to meet the physiological rigors of oviposition ( Larson, 1994; Larson, 2008b). Schweitzer et al. (2005) reported medullary bone (calcium deposition stores in the cancellous and medullary cavities of birds for use during oviposition) in the femur of MOR 1125 , a specimen previously assigned to the robust morphotype ( Larson, 2008b). Those in the latter camp have noted that large tyrannosaurids had a highly gracile body form early in ontogeny and assumed more robust features as they matured ( Currie, 2003).
In most skeletal dimensions, RSM P2523.8 is close to FMNH PR 2081 . RSM P2523.8 exceeds all known T. rex specimens (including FMNH PR 2081 ) in numerous measurements, including scapula blade width, ilium length, proximal femur width, and tibia shaft width. In additional measurement (including dentary tooth row and femur length) RSM P2523.8 notably exceeds all T. rex specimens that have been previously categorized as robust ( Larson, 2008b). However, in many length measurements (proximodistal femoral, tibial and jaw length) RSM P2523.8 is exceeded by some individuals previously categorized as gracile ( Larson, 2008b). Whether or not a robust/gracile dichotomy exists among T. rex , these comparisons indicate that RSM P2523.8 was a large and robustly proportioned individual, but likely with a shorter total hipheight and snout-vent length than other known specimens showing more elongate proportions.
Body mass (kg) 8,870
6,740
8,462 6,100 5,469 5,779 6,264 5,112 4,469 5,025 4,714 3,617 Pedal p. IV- 1 length 184 145 154 170 140? 176? 107 124?? Astragalus width 310?? 295? 280 235????? Astragalus height 328?? 280? 325?????? Fibula shaft width 62? 59 53 65 55 56? 52??? Fibula length 995 1,025 1,030 915 985 945 1,035??? 930? Tibia shaft width 184 150 160 150 155 158 170? 160? 140? Tibia length 1,140 1,166 1,140 1,060 1,095 1,065 1,150? 1,030? 1,105 910 Proximal femur width 426 399 380 370? 350 370? 270 330?? Femur circumference 590 534 580 515 495 505 520 483 460 480 469 426 Femur length 1,333 1,269 1,321 1,150? 1,350 1,280 1,232 1,210? 1,100 1,040 Ilium length 1,545 1,540 1,525?? 1,540 1,470 1,397?? 1,180? Manual p. I-1 length 98.5? 78??? 89??? 90? Scapula blade width 74. 8 73 68 65. 7?? 65????? Dentary tooth row length 595 508 585 492? 575 635 546 530??? Morph Robust Robust Robust Robust Robust Gracile Gracile Gracile Gracile??? Specimens RSM P 2523.8 CM 9380 (AMNH 973 ) FMNH PR 2081 MOR 1125 RTMP 81. 12. 1, NMC 9950 BHI 3033 MOR 555 MOR 980 RTMP 81. 6. 1 BM R8040 (AMNH 5881 View Materials MOR 009 USNM 6183 View Materials
Medullary bone structure would only be expected in a narrow window associated with eggshell formation, and there are no histological grounds for assigning RSM P2523.8 to either sex. As such, no independent support for the sexual dimorphism hypothesis is possible from the present analysis.
The extent of remodeling in RSM P2523.8 precludes direct aging using a total growth line count, as has been done for other tyrannosaurid specimens ( Erickson et al., 2004; Erickson, 2005). Nevertheless, the degree of remodeling exceeds that reported in any T. rex long bone ( Erickson et al., 2004; Horner and Padian, 2004). This suggests the animal is of at least comparable, and likely more advanced ontogenetic status than the ~28 year old, somatically mature T. rex specimen FMNH PR 2081 ( Erickson et al., 2004). Thus, the histological analysis may offer independent support for the prediction that high robustness correlates with greater maturity.
Comparisons with Other Giant Theropods
Is T. rex the largest known terrestrial carnivore ever to have lived? Based on the size distribution for described specimens of giant theropods and our new morphometric data for T. rex , RSM P2523.8 substantially exceeds the known size for all other giant theropods (1,650–6,260 kg) ( Table 2 View TABLE 2 ), including taxa previously touted as larger ( Coria and Salgado, 1995; Currie, 1996). There are, however, two important caveats that should be kept in mind when interpreting these theropod mass estimations and the conclusion based on RSM P2523.8 that T. rex was the largest terrestrial carnivore.
.
First, as the largest T. rex specimen, and presumably among the oldest, RSM P2523.8 appears to represent the chance sampling of a rare somatically mature individual. Within vertebrate paleontology, a combination of taphonomic and collection biases tend to favor the recovery of larger specimens ( Damuth, 1982; Brown et al., 2012; Brown et al., 2013). However, a bias also exists in the other direction, which makes the recovery of the largest and oldest representatives of many dinosaur taxa improbable. Erickson and Currie (2006), in a previous study surveying the size of all substantially complete T. rex specimens, showed that the taxon had a type-B1 survivorship curve (high juvenile mortality, negligible midlife mortality, and high adulthood mortality) with very few individuals reaching full adult size. As such, the recovery of older somatically mature specimens should be rare.
Because a type-B1 survivorship curve and resulting sampling bias occurs throughout the Dinosauria ( Erickson, 2005; Hone and Mallon, 2017), and because most dinosaur taxa are known from single specimens ( Wang and Dodson, 2006), Erickson (2005) and Horner et al. (2011) predicted that most known species would be found to be substantially larger than currently thought. However, T. rex is now known from over 20+ reasonably complete individuals, increasing the likelihood that one of the oldest individuals is represented. RSM P2523.8 and FMNH PR 2081 are close in estimated mass and in most skeletal dimensions, and, as both specimens are skeletally mature, they may converge upon the asymptotic mass for T. rex . The slightly greater size of RSM P2523.8 may reflect it being relatively large for its age class. In extant vertebrates, populations within an age class typically show 20% variance in body mass, and variation is even greater in taxa showing size-based sexual dimorphism ( Hanks, 1972; Navarro et al., 2005), which as previously explained, has been proposed for T. rex .
)
Although RSM P2523.8 has an estimated weight more than 40% greater than the next largest known theropod taxon, specimens of several other theropod species (including Giganotosaurus carolinii and Tyrannotitan chubutensis ) have femoral proportions indicative of body masses greater than those of most other adult T. rex specimens (compare Tables 1 and 2). As such, it is likely that further sampling of these other giant theropods, all of which are represented by fewer specimens than T. rex , may yield larger individuals that match or surpass the size of RSM P2523.8 . In the case of Giganotosaurus carolinii , a sing dentary is known that does hint at a greater maximum size ( Calvo, 2000).
Second, although RSM P2523.8 has the greatest femoral circumference among giant theropods ( Table 2 View TABLE 2 ), it does not show the greatest femoral length (the femora of Tyrannotitan chubutensis MPEF-PV 1157 are equally long and those for Giganotosaurus carolinii MUCPv-CH 1 are roughly 10% longer). Though a less reliable indicator of body mass than MFC ( Campione et al., 2014), femoral length is generally considered an isometrically stable feature across theropods. However, rather than being proportionately short-legged, the other hindlimb bones of tyrannosaurids are far more elongate than those of carcharodontosaurids ( G. carolinii has a tibia length of only 1,060 mm, compared with 1,140 mm in RSM P2523.8 ). Second, although femoral circumference has emerged as the standard for dinosaur mass estimation (and has been rigorously calibrated and tested in studies of extant animals), the method still suffers from a considerable range of error (variation from the true value of as much as 20% is generally expected) ( Campione et al., 2014). Moreover, applying the method to comparisons between tyrannosaurs and other large theropods may constitute a particular challenge.
There has been much debate over the absolute running potential (or lack thereof) of large tyrannosauroids, but however fast or slowly tyrannosauroids could run/walk, the comparative anatomical evidence indicates a relatively greater degree of limb exertion than in other large theropods ( Snively, 2000; Snively and Russell, 2003; Persons and Currie, 2016). The underlining assumptions of femoral circumference based mass estimations are that minimum femoral circumferences reflect femoral crosssectional strength, that in turn reflect the amount of stress the femur was suited to withstand, that in turn reflects how much body mass the femur needed to support. However, body mass alone is not the only factor contributing to the stress and strain applied to a limb bone. Locomotor loads from muscle, joint, and ground reaction forces are also factors. During faster and more vigorous movement, limb bones will experience greater strain than during slower and less vigorous movements. As such, when comparing the femur circumferences of two animals, it is worth considering whether the two are expected to have experienced a similar degree of limb exertion and locomotory reaction forces.
Tyrannosaurids are characterized by numerous anatomical features that suggest the group was capable of more rapid locomotion than other similarly sized theropods (including large allosauroids). These traits include an arctometatarsalian pes ( Holtz, 1995; Snively, 2000; Snively and Russell, 2003), proportionately longer distal limb bones ( Bakker and Bir, 2004; Persons and Currie, 2016), ilia with increased muscle attachment surfaces, and more rigid posterior caudal series with potential to better aid in turning and stability ( Bakker and Bir, 2004). Greater femoral circumference with the capacity to withstand greater locomotor loads, rather than simply greater body mass, merits consideration as an explanation for the discrepancies between femoral lengths and girths in T. rex and G. carolinii . Relative locomotor activity may similarly explain the extreme discrepancies in weight estimations reported for the proportionately slim-femured carcharodontosaurid Acrocanthosaurus atokensis . Notably the 3.5 tonnes femur derived estimation for mass vs. 5.5–7 tonnes based on volumetric estimation ( Bates et al., 2009).
Various authors have suggested a biomechanical ceiling that caps the body mass at which the bipedal theropod body plan can function with ecologically feasible efficacy ( Hutchinson and Garcia, 2002; Henderson and Snively, 2004; Therrien and Henderson, 2007; Hutchinson et al., 2011). As this ceiling is approached, the typical advantages of greater body mass are thought to be increasingly opposed by sizedependent restrictions on speed and agility, including bone and joint strength, and musculoskeletal torques ( Farlow et al., 1995; Hutchinson and Garcia, 2002; Hutchinson et al., 2005), biomechanical safety factors ( Farlow et al., 1995), and sheer inertia ( Henderson and Snively, 2004; Hutchinson et al., 2007). Extreme giantism in Allosauroidea and Spinosauridae may be attributable to factors that mitigate these restrictions and allowed both linages to press this ceiling, such as access to enormous, but correspondingly slow, sauropod prey in the former and the weight-supporting advantages of a semiaquatic lifestyle, in the latter. However, the evolutionary history leading to the scale of RSM P2523.8 unfolded in a purely terrestrial context. Nor were large sauropods prevalent throughout the range of T. rex , and definitive evidence of T. rex predatory attempts have been identified among the smaller, more common hadrosaurs ( DePalma et al., 2013) and purportedly ceratopsians ( Happ, 2008). Tyrannosaurid giantism therefore appears to have arisen without similar mitigating factors. As such, T. rex represents the greatest size yet known at which terrestrial predation remained viable.
CONCLUSION
Tyrannosaurus rex is arguably the most extensively studied and consequently best understood nonavian dinosaur ( Brusatte et al., 2010). Nevertheless, questions pertaining to its ontogeny and the biomechanical challenges of its great size remain pervasive. RSM P2523.8 shows the form of an elder T. rex at the terminal extreme of both growth and maturation. Despite its formidable size, T. rex was evidently not invulnerable, and the relative scarcity of other equally large and mature T. rex specimens suggests a high rate of mortality within the species below an 8-tonne threshold. Similarly, the various skeletal pathologies of RSM P2523.8 record the numerous severe injuries suffered over the course of its violent and unusually long life.







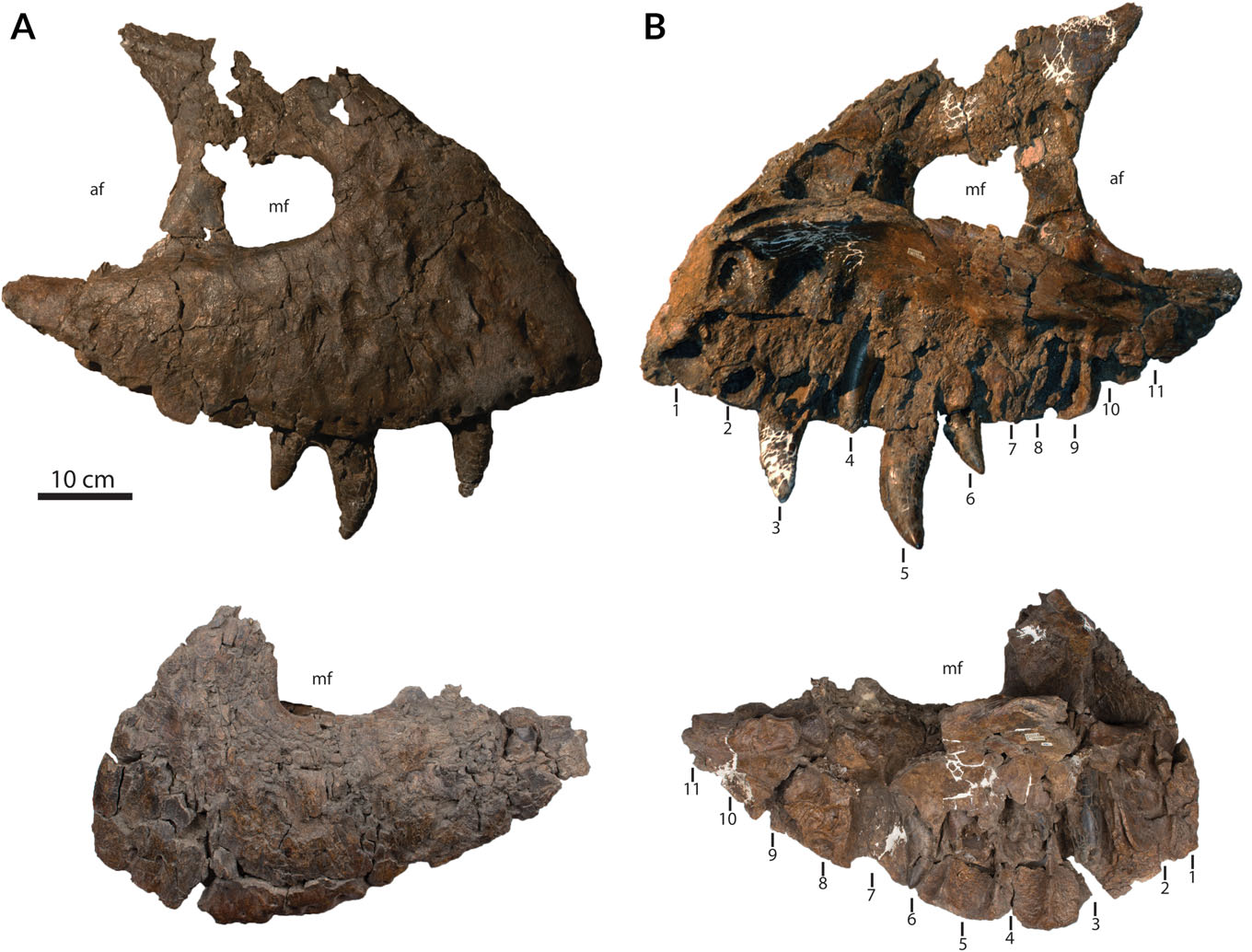





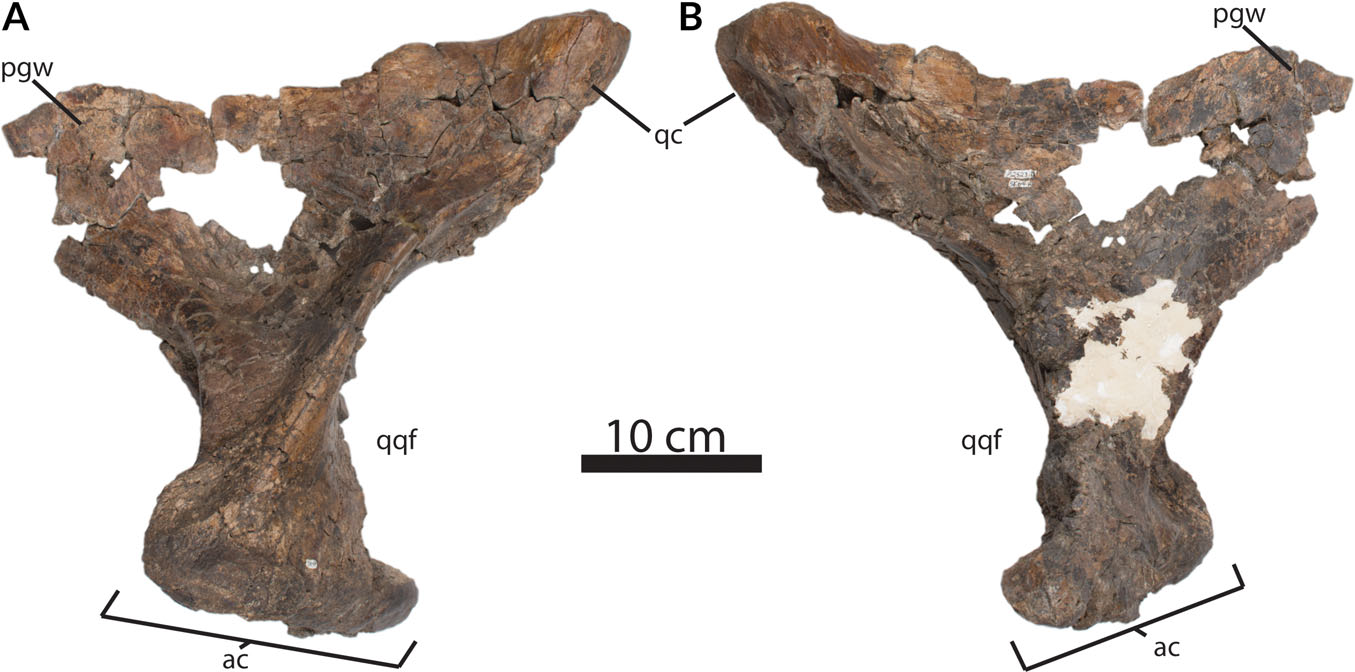


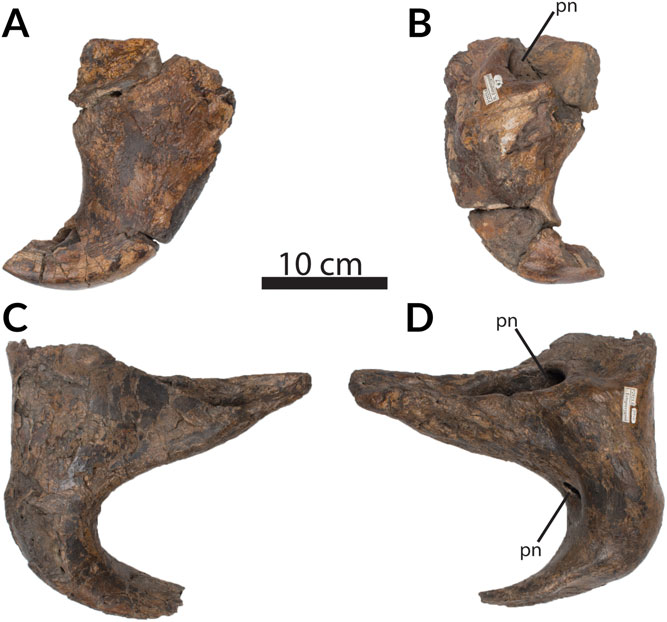


TABLE 2. Comparison of femoral dimensions and estimated body mass of the largest theropods ±25% (mass estimation based on the formula of Campione et al. (2014))
| Species | Specimens | Femur length (mm) | Femur circumference (mm) | Body mass (kg) |
|---|---|---|---|---|
| Tyrannosaurus rex | RSM P2523.8 | 1,330 | 590 | 8,870 |
| Acrocanthosaurus atokensis | NCSM 14345 | 1,277 | 425 | 3,590 |
| Chilantaisaurus tashuikouensis | IVPP V.2884 | 1,190 | 430 | 3,710 |
| Deinocheirus mirificus | MPC KID447 | 1,300 | 520 | 6,260 |
| Giganotosaurus carolinii | MUCPv-CH 1 | 1,365 | 520 | 6,260 |
| Mapusaurus rosei | PVPH.108.44 | 1,170 | 435 | 3,830 |
| Mapusaurus rosei | PVPH.108.203 | 1,205 | 415 | 3,370 |
| Mapusaurus rosei | PVPH.108.233 | 1,115 | 420 | 3,480 |
| Saurophaganax maximus | OMNH 01708 | 1,130 | 435 | 3,830 |
| Spinosaurus aegyptiacus | Ibrahim et al. (2014) | 830 | 320 | 1,645 |
| Suchomimus tenerensis | MNN GDF 500 | 1,080 | 410 | 3,260 |
| Tyrannotitan chubutensis | MPEF-PV 1156 | 1,265 | 520 | 6,260 |
| Tyrannotitan chubutensis | MPEF-PV 1157 | 1,330 | 505 | 5,780 |
Note. There are a great number of problematic assumptions in the mass estimate of Spinosaurus aegyptiacus , see main text for details.
| RSM |
RSM |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |