
Ruellia L .
|
publication ID |
https://doi.org/10.5281/zenodo.11066989 |
|
DOI |
https://doi.org/10.5281/zenodo.11104237 |
|
persistent identifier |
https://treatment.plazi.org/id/0F7687E0-1B68-FF9A-D7FA-FD33A347033C |
|
treatment provided by |
Felipe |
|
scientific name |
Ruellia L . |
| status |
|
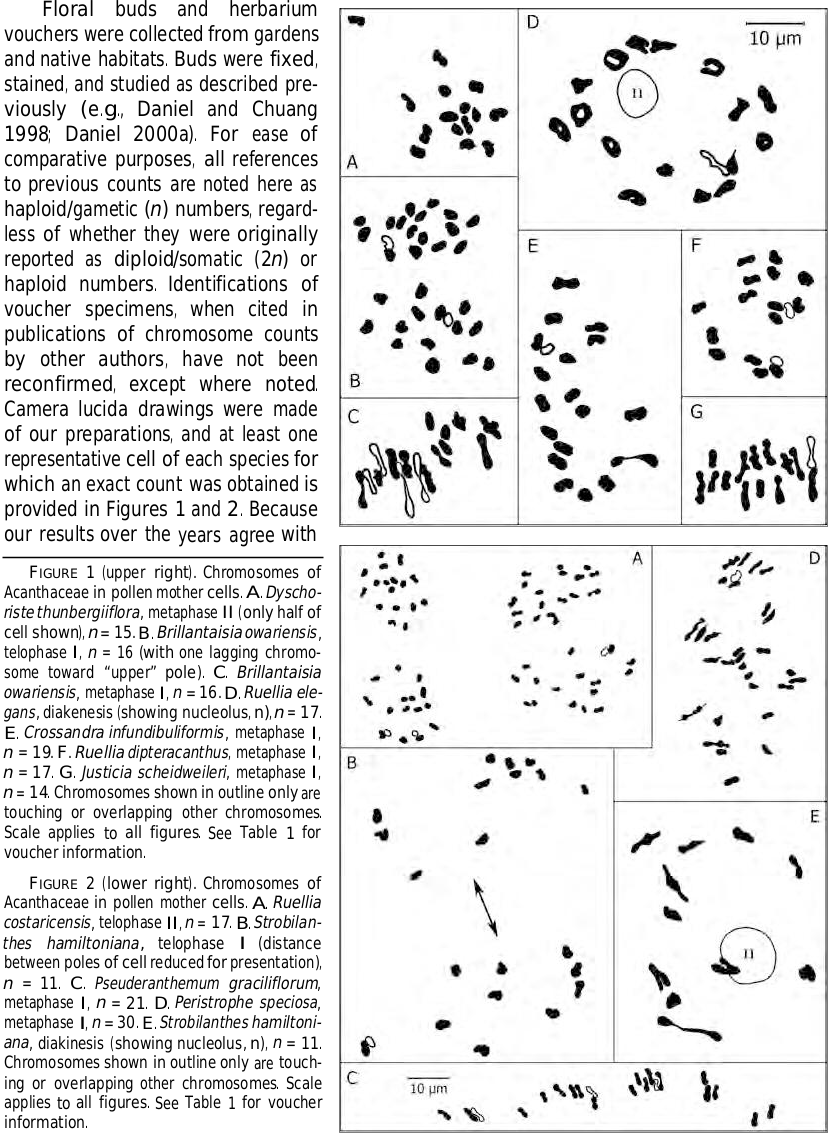
— Ruellia (Ruellieae: Ruelliinae) consists of about 350 species worldwide, with most of them occurring in the Western Hemisphere. The chromosome number n = 17 is common to at least 43 species occurring throughout its range in the New World (i.e., North America, Central America, South America, and the West Indies). Our counts of n = 17 (or ca. 17) for four Neotropical species confirm this number for them. Chromosomes of R. costaricensis (Oerst.) Tripp & McDade ( Fig. 2A View FIGURE ), a species of Nicaragua, Costa Rica, and Panama, were previously counted by Daniel (2000a, as Blechum costaricense Oerst. ) for this rarely cultivated species. Our count for R. dipteracanthus (Nees) Hemsl. ( Fig. 1F View FIGURE ), presumably native to Mexico, agrees with previous counts by Grant (1955, as R. squarrosa (Fenzl) Schaffnit , and as “ R. fluviatilis Leonard ,” nom. ined., voucher at US seen). The counts for two Brazilian species, R. elegans Poir. ( Fig. 1D View FIGURE ) and R. makoyana Closon confirm previous counts for them by De (1966, as R. formosa Andr. ) and by Grant (1955), respectively. The taxonomic relationships of R. makoyana to its morphologically similar congeners R. devosiana E. Morren (also n = 17; e.g., Daniel and Chuang 1998) and R. portellae Hook. f. , all attributed to Brazil, are worthy of additional study. The only other chromosome numbers reported for species of Ruellia native in the New World (discounting two counts of n = 18 by Sugiura (1939) because no vouchers were cited, identities of the species are questionable because no authors of names were given and one of the names applies to multiple species, and the counts reported differ from all subsequent reports for those species) are n = 16 for R. tuberosa L. (the majority of counts for this species is n = 17, see summaries of counts in Daniel 2000b) and n = 24 for Brazilian R. macrantha (Nees) Mart. ex B.D. Jacks. ( Daniel & Chuang 1998).
In the Old World, there is a single report of n = 17 for R. prostrata Poir. ( Sarkar et al. 1980); however, other numbers have also been reported for this species (e.g., n = 16, Subramanian and Govindarajan 1980; n = 24, Daniel and Chuang 1998), and n = 22 has been reported for it several times (e.g., De 1966; Kaur 1966; Govindarajan and Subramanian 1983; Saggoo and Bir 1983). All of the counts for this African and Asian species apparently pertain to plants from India. Other counts for Paleotropical species of Ruellia consist of n = 16 for African R. cordata Thunb. (Rao and Mwasumbi 1981) and for Asian R. patula Jacq. (e.g., Baquar et al. 1966; Rao and Mwasumbi 1981; Subramanian and Govindarajan 1980; Govindarajan and Subramanian 1983) and n = 12 for Asian/Malesian R. repens L. ( Daniel 2000b).
Based on the small sampling of species in the Old World, the diversity of chromosome numbers there appears to be greater than that for taxa in the New World. Tripp (2007) indicated that taxa in the New World appear to be monophyletic and derived from African stock. Radiation from a single introduction in the New World might explain the widespread occurrence of n = 17 among diverse taxa there. Numbers common to both Neotropical and Paleotropical species are n = 16, 17, and 24. At least the latter suggests potential polyploidy in the genus. Meiotic complements of n = 16 are common elsewhere among Ruellieae (see discussions herein for Brillantaisia , Dyschoriste , and Strobilanthes ). Based on numbers reported to date, x = 12 or 16 would appear to be potential basic numbers in Ruellia , from which numbers of n = 17 and 24 may have arisen via dysploidy and polyploidy. If counts of both n = 16 and 17 for R. tuberosa are accurate, then dysploidy likely also occurs in some species. Although not currently known for any species of the genus, an ancestral basic number of x = 8 for either Ruellia (or Ruellieae) might account for lineages with n = 16 and 24. Given a potential ancestral basic number of x = 7 for the family (e.g., Piovano and Bernardello 1991; Daniel 2000a), such an hypothesis has some appeal. No chromosome numbers have been reported for any of the other genera of Ruelliinae (i.e., Acanthopale C.B. Clarke , Dischistocalyx T. Anderson ex Benth. , and Satanocrater Schweinf. ; Tripp et al. 2013). Counts for these genera and additional counts for species of Ruellia in the Old World are needed to better formulate a potential ancestral basic number for and to understand patterns of evolution and migration in the genus. Counts are especially desired for species native to Madagascar and Australia, from which regions no chromosome numbers have been reported for any Ruelliinae.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |