
Acantorna tahoma, Lehnert & Stone, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4755.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:FFDB122C-E34B-486A-AC99-88B240AAC4DD |
|
DOI |
https://doi.org/10.5281/zenodo.3730245 |
|
persistent identifier |
https://treatment.plazi.org/id/0E138793-FF95-AF20-FF2D-F6990C95A763 |
|
treatment provided by |
Plazi |
|
scientific name |
Acantorna tahoma |
| status |
sp. nov. |
Acantorna tahoma View in CoL n. sp.
( Figs 2 View FIGURE 2 & 3 View FIGURE 3 )
Material examined. Holotype ZSM 20200044 View Materials , intact specimen, frozen then dried, collected by Bill Flerx with a research survey bottom trawl from the FV Sea Storm; 4 August 2012, 229 m depth, haul station 218, west of Tahoma Reef , western Aleutian Islands (52° 09.84’ N, 175° 13.41’ E). Water temperature = 3.8 °C. GoogleMaps
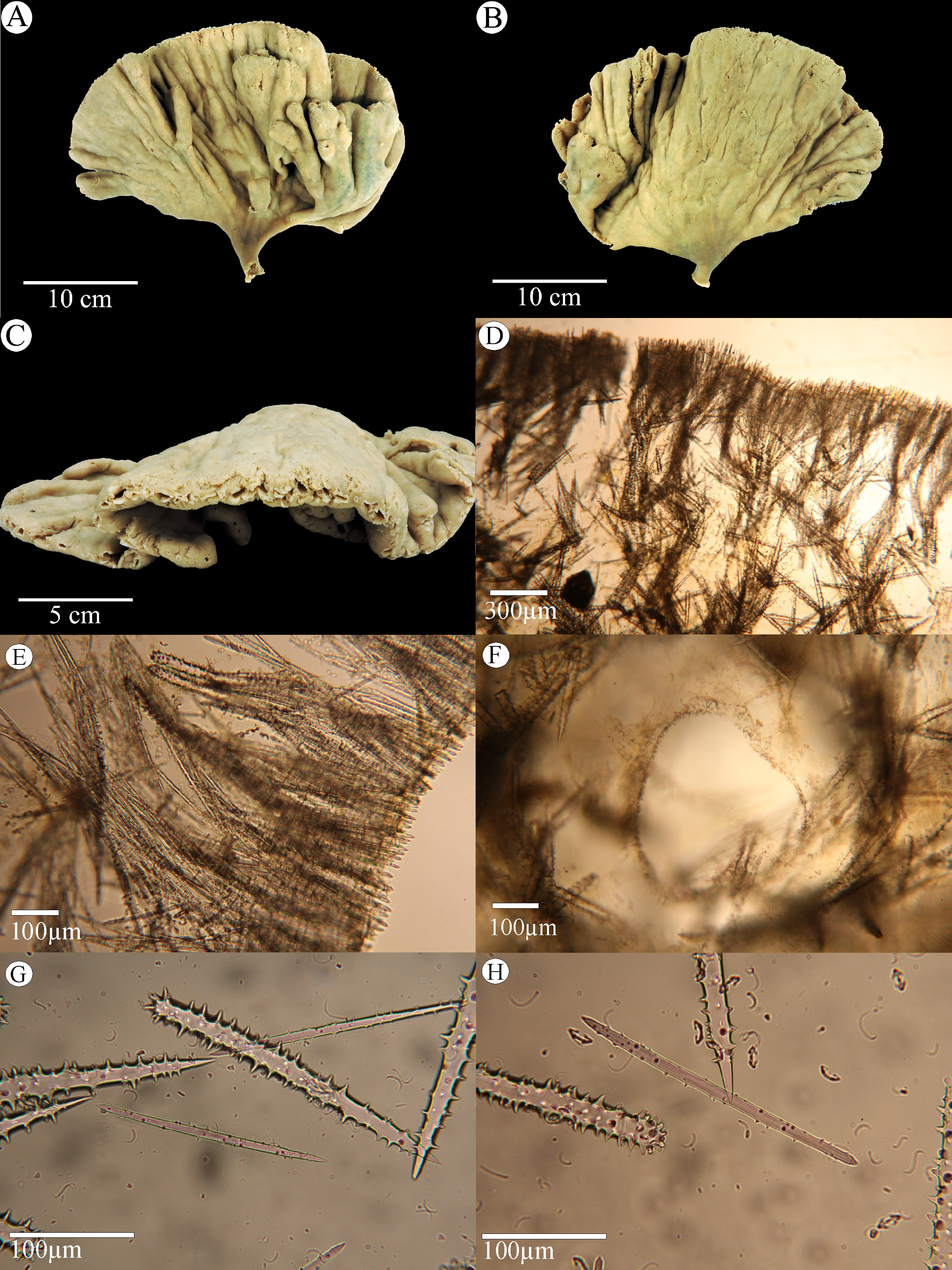
Description. The holotype is a stalked, ochre coloured, flabellate sponge ( Figs. 2 View FIGURE 2 A-C). The stalk is 17 mm long and 17 mm in maximum diameter and then widens gradually into a fan. The fan is undulating. Mainly radial incisions are visible on the surface of the fan, running to the top. The sponge reaches a maximum height of 28.5 cm and width of 29 cm. The thickness of the fan is up to 1.2 cm. The surface is optically smooth, velvety. Circular oscules with a thin, slightly elevated rim, 1–3 mm in diameter are restricted to the upper rim of the fan ( Fig. 2C View FIGURE 2 ). Consistency of the stalk is wiry and incompressible and becomes more elastic towards the top. The fan is slightly elastic and tough. It has five side-branches at various heights and of different dimensions, the smallest is 2.7 x 0.8 x 0.5 cm, the second is 2.9 x 1.6 x 0.7 cm. The latter has an incision at the top. The three larger side branches are smaller, stalked fans and the two largest „side-fans“ anastomose at the margins. The largest measuring 11 x 9 x 0.9 cm, with a row of oscules at the top ( Fig. 2C View FIGURE 2 ).
Skeletal architecture. The choanosome is a confusedly plumose arrangement of acanthostyles ( Figs. 2 View FIGURE 2 D-E), tracts are not well defined but with a clearly vertical denser portion of spiculated tracts separated by lightly spiculated areas in between those tracts are restricted to the choanosome, and acantornotes, arcuate isochelae and sigmas occuring throughout the sponge. Spicule tracts of acantornotes start in a subectosomal zone, 0.6–2 mm below the surface ( Figs. 2 View FIGURE 2 D-E), terminating in dense surface brushes forming a palisade ( Figs. 2 View FIGURE 2 E-F). Choanosomal canals are lined by a thin veneer of microscleres ( Fig. 2F View FIGURE 2 ).
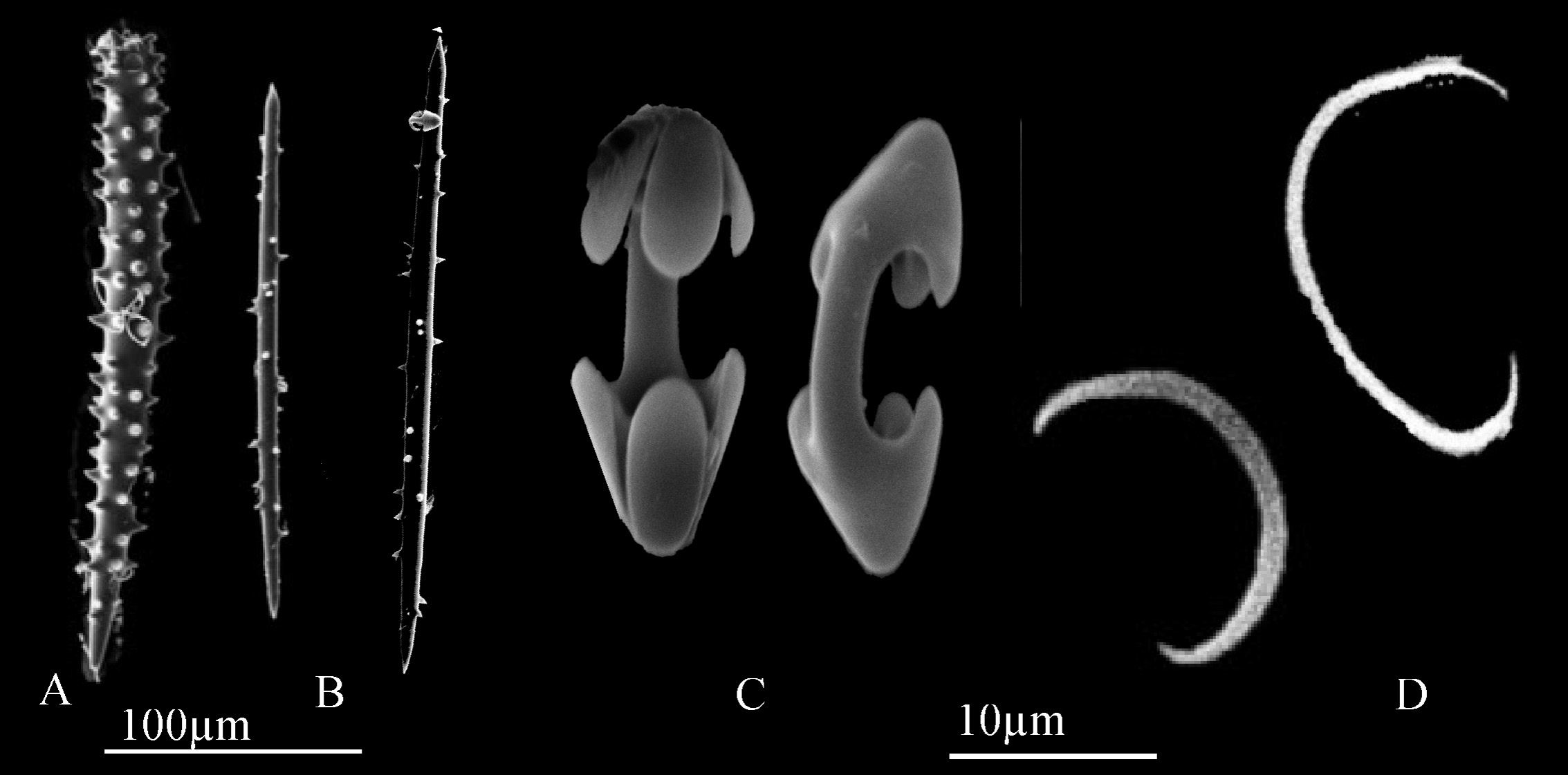
Spicules. Choanosomal acanthostyles ( Figs. 2G View FIGURE 2 , 3A View FIGURE 3 ), 240–311–329 x 24–31– 36 µm; choanosomal and ectosomal acantornotes ( Figs. 2H View FIGURE 2 , 3B View FIGURE 3 ); 155–191–283 x 8–9– 10 µm, often with unequal ends; arcuate isochelae ( Fig. 3C View FIGURE 3 ), 18–21– 24 µm; abundant thin sigmas ( Fig. 3D View FIGURE 3 ), 12–17– 24 µm.
Etymology: The species name tahoma refers to the type locality near Tahoma Reef in the western Aleutian Islands.
Discussion. This species is clearly a Poecilosclerida due to the presence of chelae, and with the possession of arcuate chelae and the lack of toxas placing it within the now-abandoned suborder Myxillina (polyphyletic; Morrow & Cardenas 2015). As families with arcuate chelae were united in Myxillina it would have been sufficient to compare this new taxon only to families assigned to the group, but we also compare it with Microcionidae because they share diactinal ectosomal spicules in some rare cases, and the arrangement of subectosomal spicules. In agreement with the three distinct skeletal zones of the former Myxillina we can also distinguish three different zones in the sponge described here. The ectosomal brushes of tornotes conform to zone 1 with the only difference that acantornotes replace the smooth tornotes, the subectosomal zone, or zone 2 in our sponge, is not represented by a special type of spicule but replaced by spicule tracts of the same acantornotes, but this is not exceptional as reduction in skeletal components was also known to occur in taxa assigned to the Myxillina . The choanosome conforming to zone 3 deviates from the Myxillina skeletal plan as it contains all spicule types in confusedly plumose arrangement, not known from other Myxillina taxa. Tornotes were known as smooth or terminally microspined, whereas tornotes which are completely acanthose are so far unique. The combination of a choanosome with a confusedly plumose arrangement of all occurring spicules types and the occurrence of the previously unknown acantornotes support the erection of a new genus.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |